阿 Q 為了救回被黑暗料理界綁架的父親,與小當家一行人前往樓麟艦,接受黑暗料理界所下的戰帖,展開四回合的宴席料理戰。第一回合的對決主題是「湯」,自告奮勇擔任先鋒的阿 Q,以「彈跳甲魚湯」這道料理,漂亮地戰勝錦毛虎駱可的頂級雞湯。
彈跳甲魚湯因為凝固成魚凍不會灑出來,評審們紛紛驚嘆這是需要嚼的湯?為什麼會變成這樣呢[1]?

甲魚即是鱉,在亞洲經濟、營養價值皆高
甲魚其實就是鱉,又名圓魚、團魚或王八,是一種高經濟價值的水產養殖物種,其肉、 血、膽、脂肪,甚至是甲殼皆可以被利用。
中華鱉(Pelodiscus sinensis 或 Trionyx sinensis)目前是亞洲最普遍的品種,主要分布於中國大陸,其次為日本、臺灣、韓國、馬來西亞和泰國等,估計阿 Q 就是拿中華鱉去料理的。
鱉在部分亞洲國家已經有很長的食用歷史,大多是當作食物或傳統藥材使用,中國明朝《本草綱目》中記載「鱉甲乃厥陰肝經血分之藥,肝主血也。試常思之,龜鱉之屬,功各有所主。鱉色青入肝,故所主者,瘧勞寒熱,痃瘕驚癇,經水癰腫陰瘡,緣厥陰血分之病也。」,說明鱉具有滋陰潛陽、清熱消淤等多種漢方保健效果。
當然,鱉肉營養價值的確很高,其肉質細嫩、味道鮮美、營養豐富,是蛋白質、膠原蛋白和礦物質良好的來源。成分包含 20 種必需胺基酸、EPA、DHA 和其它不飽和脂肪酸,豐富的微量元素包含鋅、錳、鐵、銅、鈣、磷、維生素 A、B1、B2 及 D 等[2]。

凝膠性是蛋白質重要的功能特性之一
這就不得不說明,蛋白質的功能特性。
蛋白質是細胞重要的組成分,人類從食物中攝取蛋白質,不外乎就是為了補充營養上所需的必需胺基酸。
但是蛋白質在食品的應用上,具有一些功能特性,例如凝膠性(gelation)、黏彈性(viscoelasticity)、乳化性(emulsification)或起泡性(foaming)等,這些特性在生活中隨處可見,舉例來說:
做麵包時,將麵粉(小麥蛋白)加水攪打成具有彈性的麵團,就是黏彈性;製作蛋黃醬時,將蛋黃跟醋酸、沙拉油這兩樣油水不溶的材料均勻混合在一起,就是乳化性;做蛋糕時,將蛋白打發成乾性發泡的蛋白霜,就是起泡性。
而彈跳甲魚湯所牽涉到的蛋白質功能特性,就是凝膠性。
蛋白質凝膠,是變性的蛋白質分子在一些作用力包括氫鍵、疏水交互作用或靜電斥力交互作用等影響下,蛋白質分子互相聚集、吸引或排斥達成平衡,以至於形成能保持大量水分的高度有序之三度網狀結構,是一個動態的過程[3]。
彈跳甲魚湯會凝膠是因為明膠
鱉所富含的「膠原蛋白」是彈跳甲魚湯「凝膠」的關鍵,鱉肉經過長時間的熬煮,膠原蛋白轉變成「明膠」,靜置冷卻後便凝固成果凍狀,是一種肉凍料理[4],像豬肉凍、水晶肴肉都是類似的料理。
P.S. 鱉的脂肪也有可能影響到這道料理的形成,但並非主因,本文為避免複雜,就不再探討脂肪的食品化學。
明膠(gelatin)為膠原蛋白(collagen)熱變性後的產物,大量存在脊椎動物中,動物性蛋白質約有 30% 是由膠原蛋白構成,含量會隨著年紀及季節的不同而改變,存在腱、皮膚、骨、血管、結締組織等[5]。
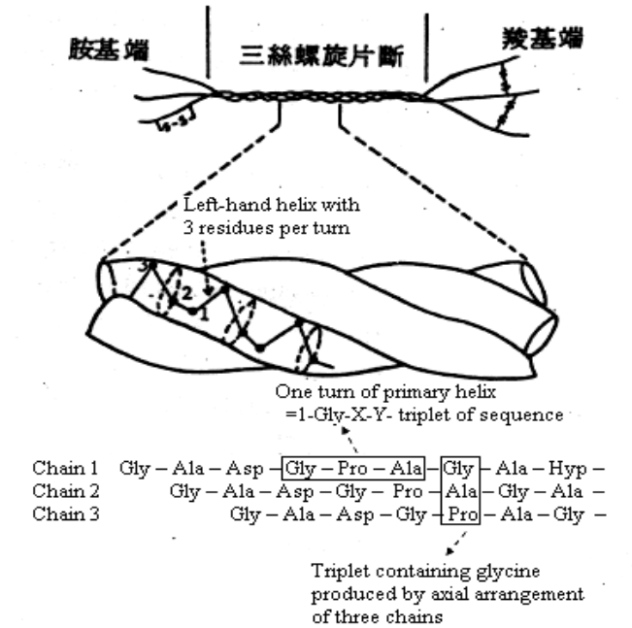
膠原蛋白係由三條多胜肽鏈 (polypeptide chain) 相互纏繞而成的三股螺旋結構,多胜肽鏈上以甘胺酸(glycine, Gly)為最豐富的胺基酸,約佔 33%,其次為 13% 羥脯胺酸(hydroxproline, Hyp)、 12% 脯胺酸(proline, Pro)、11% 丙胺酸(alanine, Ala),以及稀有之 1% 羥離胺酸(hydrolysine,Hyl)等[4]。
這三條多胜肽鏈相互以氫鍵緊密纏繞連結,如同堅韌的繩索一般,所以肉的膠原蛋白含量愈多時,肉質就較硬。不過當以攝氏 40 度以上加熱時,膠原蛋白分子間的氫鍵被打斷,破壞三股螺旋結構,即轉化為水溶性之明膠[5]。
而明膠冷卻後,分子間再度以氫鍵鍵結而「凝膠」形成果凍狀,這個凝膠只要再加熱破壞分子間氫鍵,又會恢復成流動狀,是熱可逆反應。
吃膠原蛋白不能補膠原蛋白?
膠原蛋白是維持人體肌膚彈性的要素之一,隨著年紀增長,皮膚真皮層的膠原蛋白含量減少,令愛美女性在意的皺紋就會出現。
那喝彈跳甲魚湯,或是香甜滑溜的銀耳湯,可以幫助我們補充膠原蛋白嗎?
事實上,膠原蛋白本來就是人體可自行合成的物質,以不同的形式存在於皮膚、骨骼、軟骨、韌帶、肌腱、血管壁和結締組織等部位,並不需要額外補充。
不管是從食物攝取,或是吃膠原蛋白補充品,一樣都會經過人體消化作用,變成小分子的胺基酸,未必能在體內重新合成膠原蛋白[7]。
且豬腳、雞爪要是吃過量,反而容易攝取到過多的油脂與熱量,不但沒達到美容效果,還導致肥胖。
銀耳是植物,植物不含膠原蛋白
另外,許多女性喜愛的銀耳湯,其實是不含膠原蛋白的。
許多人以為,滑溜溜的銀耳含大量的膠原蛋白,事實上,膠原蛋白多存在於動物中,植物不含膠原蛋白。銀耳屬於蔬菜類中的菇類,那滑溜的口感是來自於「多醣體」。
銀耳含有豐富的多醣體,屬於水溶性纖維,能吸收水分、增加飽足感、延緩血糖上升,同時也是腸道好菌的食物來源。

只要均衡飲食、適量攝取蛋白質、少吃高油炸食物、補充維生素 C 或維生素 E 等抗氧化物質,並且生活作息正常,就能減少體內膠原蛋白的流失,達到維持肌膚彈性的目的[8]。
參考資料
- Muse木棉花,2021。中華一番(舊版小當家) 第36話【阿Q特製!冷卻煮凝湯】。
- 陳思宇,2008。甲魚 (中華鱉) 蛋理化特性之探討。輔仁大學食品科學系碩士論文。新北。
- 劉展冏、韓建國、劉冠汝、李嘉展、虞積凱、孫芳明、蘇敏昇、馮惠萍、謝秋蘭、饒家麟、梁弘人、林聖敦、江伯源、李政達、盧更煌、周志輝,2020。最新食品化學(最新修訂版)。於陳建元修編,顏國欽總校閱。臺中市:華格那出版有限公司。
- 唐嘉憶,2006。明膠作為配料利用添加於肉凍製品之研究。國立屏東科技大學食品科學系碩士學位論文。屏東。
- 黃鈺茹、蕭泉源,2011。不同水生生物來源所得之膠原蛋白物理與生物化學相關特性。海大漁推 41,17-52。
- Woodhead-Gallowy J. 1980. Collagen: the Anatomy of a Protein. Journal of Anatomy 132: 3 433-437.
- 衛生福利部食品藥物管理署,2015。市面上有許多含膠原蛋白的保養品,請問真的能有效維持肌膚彈性嗎?。食藥闢謠專區。
- 衛生福利部食品藥物管理署,2022。滑溜溜的銀耳湯,真的能補到妳的膠原蛋白嗎?。藥物食品安全週報 876:7。