
還記得小時候跟著媽媽去買菜,被問到今晚想吃什麼菜,常常只能望著架上一片綠意盎然但卻不知眾菜何名的心情,最熟悉、也最會分辨的青菜通常只有,白菜、高麗菜、花椰菜。
不曉得現在的小朋友們是否和當年的我一樣?
直到長大後多讀了一點書,這才驚覺——原來,白菜、高麗菜、花椰菜,竟然是親戚?!
風靡歐亞、愛冷的蔬菜大家族——蕓薹屬
這一家子不僅家大業大,還可說是一門忠烈,煎、煮、炒、炸、燉、烤、紅燒、生食、熱飲,無論東、西方料理都獻身無數,除了可以吃,還拿來榨油、劈砍、作拐杖,怪不得美國人為了感念這一家子的貢獻,還特地訂了一個「全國甘藍日」註1以資紀念。
這一大家族,有個文謅謅的名字——「蕓薹屬」(Brassicas)是也!

在自然狀況下,這家的成員都有類似的生命模式,通常這類植物都比較喜愛冷涼的生長環境,會在秋冬季時努力生長,直到春天來臨、氣溫回暖才開花結籽,由於這樣的生長模式會跨越人類的冬季至春季陪人類過年,所以也被稱為「二年生植物」(biennial plant)。
植物學家一直很想建立蕓薹屬家族的族譜,但最早的祖先實在難以考究,只知道大約是原生於歐洲及中東地帶,並在中國西北部、地中海地區及西歐溫帶地區獨立或共同演化成現在這樣子。
目前我們能夠確定的是,依照「染色體數目」來區分可以把蕓薹屬家族再分成三大分支,分別是:
- 白菜群,有 10 條染色體
- 甘藍群,有 8 條染色體
- 黑芥群,有 9 條染色體
其中的白菜群主要在亞洲地區演化,而甘藍群則主要在歐洲育成,並各自成為兩大洲人民重要的蔬菜來源。

油菜、泡菜、酸菜,大家都是好「白菜」
白菜群的祖先,就是「蕓薹」(Brassica rapa),至今在中東地區還找得到野生的蕓薹,很早就有人在種,原先是拿來吃,後來發現蕓薹的種子含油量高,只要經過簡單的擠壓就可以榨出油來,就有人開始專門種來榨油。
經過幾千年來人為的選擇,要吃葉子的就選葉子大又不苦的,就選出了今天的白菜,要拿來榨油的就選種子產量高油份又多的,變成了今日的油菜。
無論是白菜或油菜,其實都是一個總稱而已。以白菜為例,白菜又可分成小白菜、大白菜、結球白菜、塌棵菜、京水菜,在亞洲各地都是很具代表性的蔬菜。
其中特別值得一提的是結球白菜,在中國的山東白菜做成酸菜酸香迷人、日本則做成漬白菜、德國酸白菜拿來配豬腳無人不知,在韓國被做成韓式泡菜被視為重要文化資產,我們實在不知道到底是誰先拿結球白菜來發酵的,但假如少了醃白菜,各國料理遜色得絕對不只三分啊!

抓住歐洲人的胃!西方甘藍群好吃又好養
甘藍群的祖先呢,則是一種叫做「野生甘藍」(Brassica oleracea)的植物,現在也還能在地中海、英國及法國部分富含石灰質的海岸上見到。
這種植物生性耐鹽耐鹼耐貧瘠,長有厚厚的葉子和又粗又韌的根,使它可以在缺水又艱困的海岸生存。
雖然不太懂一開始拿它來吃的歐洲人在想什麼啦,但一樣經過人為選擇且歐洲人更具創意,根、莖、葉、花都有人用!
愛吃葉子的就選出了今日吃葉的羽衣甘藍、中國芥藍、高麗菜,葉子不好吃但好看的就選出葉牡丹拿來觀賞;吃莖的就選出球莖甘藍(大頭菜或稱苤蘭);崇尚美食的法國人則選出了花椰菜與青花菜註2。

其中最有創意的非英國人莫屬了,英屬澤西島上的人們種有一種奇特的甘藍品種,它的莖長的又長又硬,最高紀錄據說可長到 5 公尺,至於它的用途呢,則是用來作拐杖的,也真是夠奇葩的了。
什麼叫做「野生」的甘藍?
順道一提,為什麼我們要叫這類植物的祖先叫做「野生」甘藍呢?野生、不野生,有什麼差別?
「野生」甘藍代表的是:它還沒有被人類馴化(domestication)!也就是說,野生甘藍的生長與生殖尚未被人類所支配、掌控。
那我們該怎麼判斷一種植物已經受到「馴化」?
在沒有人為干擾的情況下,野生甘藍需要傳播種子時,一旦它的種莢成熟後,種莢就會「自動裂開」,並將種子傳出去。
然而,當其後代漸漸被人類馴化之後,由於人類需要自己動手收穫種子來使用或種植,因此就會盡量挑選成熟之後「種莢不容易裂開」的植物。在植物育種學上,這個特徵會使用不脱粒性(non-shattering)來描述。

除此之外,被馴化的另一個指標是「對人類有用的特性越來越強化」,其中花椰菜就是其中一個極端的例子。
花椰菜能長出異常膨脹而大量的花原基,但其中大部分都無法正常分化成花朵,假如不考慮人類食用的功能,那麼這種特性完全就是浪、費、能、量,根本不可能在自然天擇環境下長期生存。
野生甘藍到底是何時被人類馴化?這仍是個未解之謎,但早在古希臘羅馬時代,就有人宣稱他家的甘藍不僅好吃、還有藥效,吃了可以減緩頭痛、解宿醉,因此可能早在羅馬時代,甘藍就已經是常見的盤中飧。
黑芥群雜交後也是很好吃的!
至於黑芥群的祖先「黑芥」(Brassica nigra)就更少人認識了,至今仍然沒有人為了商業生產而種植。
從演化上來看,黑芥群與白菜、甘藍群的關係也要更遠一些,但黑芥與白菜雜交產生的芥菜卻是中國菜系裡不可少的一種食材,我實在很能想像客家菜裡少了酸菜、福菜、梅乾菜會是怎樣!
找出蕓薹屬錯綜複雜的關係:禹氏三角
我們現在能對蕓薹屬家族的關係稍微理出個頭緒,必須感謝韓國的現代農業之父——禹長春(Woo Jang-Choon, 1898-1959)。
透過對蕓苔屬植物染色體數目及形態分析的研究,禹長春提出一個理論:那就是在所有的蕓苔屬植物裡,凡是染色體數相同的都是同一個「種」,而其中二倍體的白菜群、甘藍群、黑芥群可以形成三個頂點,頂點兩端的物種分別雜交就產生了四倍體的芥菜、衣索比亞芥菜及現代的油菜。
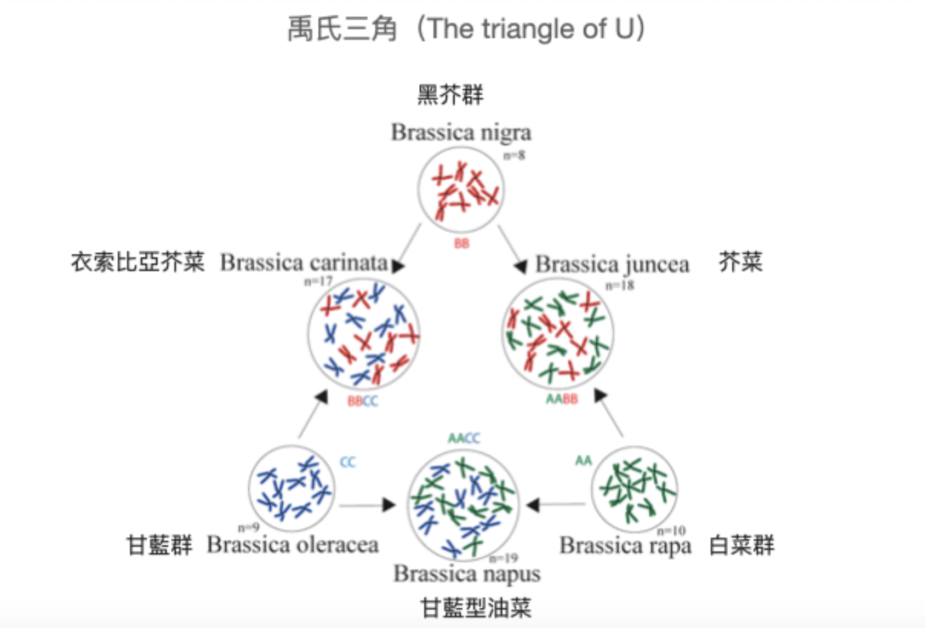
後來我們將這個理論就稱為「禹氏三角」(The triangle of U),禹長春並在1935年成功使用野生甘藍與蕓薹雜交產生非常類似現代油菜的品種,重現了現代油菜的演化過程。
現代仍然有許多育種家使用這個理論培育新品種,例如日本就利用甘藍與小白菜雜交產生千寶菜,又用青花菜與芥蘭菜雜交產生青花筍,我們的臺南農業改良場在 2018 年也利用青江菜與耐熱的油菜雜交,推出較適應台灣夏季炎熱氣候的深綠色蔬菜品種。
看來芸薹屬家族的關係只可能更亂更複雜,但也讓人期待,未來一定會有更多奇妙又好吃的蔬菜會從這個家族中脫穎而出!
備註
- 美國的全國甘藍日訂於 2 月 17 日,訂定者未知。
- 花椰菜與青花菜所食用的部分其實並不是花朵,花椰菜吃的是尚未分化完全的花原基,而青花菜又名西蘭花,吃的是尚未開花的花苞。
參考文獻
- 謝明憲、許涵鈞、王仕賢 2011 十字花科蕓薹屬蔬菜育種趨勢與生技應用概況 農業生技產 業季刊 25: 46-52。
- Maggioni, L., Bothmer, R. von, Poulsen, G. & Branca, F. (2010). Origin and domestication of cole crops (Brassica oleracea L.): linguistic and literary considerations. Economic Botany, vol. 64 (2), pp. 109–123.
- Jules, Janick (2009). Plant Breeding Reviews. 31. Wiley. p. 56. ISBN 978-0-470-38762-7.











































{kind=link}
{kind=link}