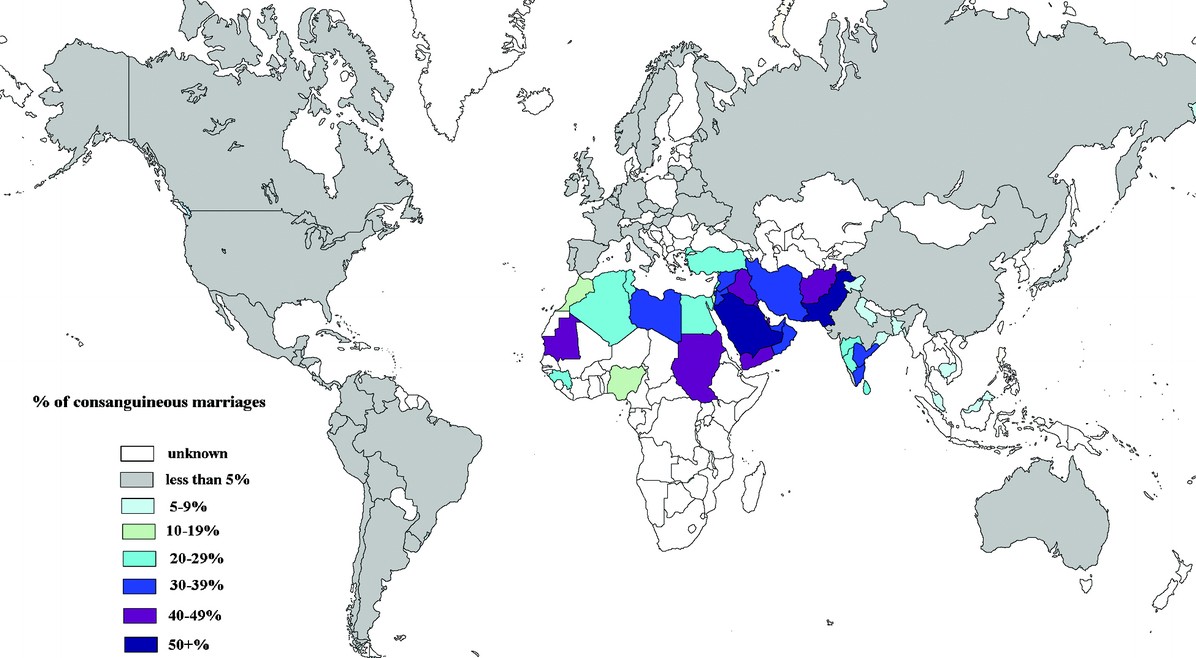
現代台灣社會中,像是堂兄弟姊妹之間的近親結婚,直接受到法律禁止。不過台灣法律的標準並非舉世通用,當今世上許多人的父母,可謂血緣上的親上加親。
近親結婚與近親繁殖,是人類的「常態」嗎?近年蓬勃發展的古代 DNA 研究,讓我們有機會深入探索這些問題。
每個人的遺傳組成都大同小異,兩個人的血緣關係愈近,彼此 DNA 的差異愈小。例如街上隨便找兩位台灣人,即使非親非故,台灣人彼此間的血緣差異,要比台灣人與非洲人更小。
一個人的基因組,源自父母各一半。例如第十一號染色體,各有一條來自父母。父母間的血緣關係愈近,小孩的一對染色體之間也愈相似;因此,要判斷一個人的父母是否為近親,不用知道兩人各自的遺傳訊息,只需要小孩的基因組。
也就是說,假如有幸獲得一位三萬年前古人的基因組,只要這個古代基因組殘留的 DNA 訊息夠多,即使完全缺乏其餘的考古脈絡,我們也能判斷他父母的血緣親疏。
最近十年來,各路科學家獲得愈來愈多古代基因組。儘管數量有限,不過目前應該足以做出初步推論:近親繁殖不是智人的天性。
尼安德塔人的父親母親,親上加親?
討論智人以前,先來看看我們的近親尼安德塔人。兩群人的祖先超過 50 萬年前分家後,各自在非洲與歐洲發展,總人口應該都不多。
這兒要先澄清一個概念:「族群人口少」和「近親繁殖」是兩回事。即使全體族群只有兩千人,整群人的遺傳變異加起來很有限,只要每一次配對時刻意選擇,依然能完全避免近親繁殖。相對地,就算總共有 20 萬人,還是有機會大量近親生寶寶。
重現尼安德塔人 DNA 是智人的重大成就,可惜目前為止累積的基因組樣本很少,只有 30 人左右,分散在不同時間點,廣大的地理範圍。
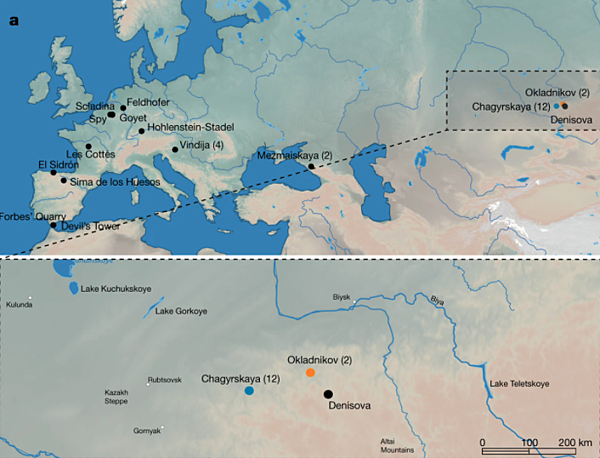
如今了解最透徹的尼安德塔人,位於中亞的 Chagyrskaya 洞穴(現今的俄羅斯南部,知名的丹尼索瓦洞穴在附近),估計年代為 5 萬多年。這群人中有 8 位的遺傳訊息比較齊全,比對得知,所有人的父母都是近親!
尼安德塔人主要住在歐洲,中亞的人口極少。近親生寶寶如此普遍,或許是由於能選擇的對象有限。然而也有可能,這就是尼安德塔人一般的習慣。也許尼安德塔人不會刻意避免近親繁殖,不過程度如何並不清楚。
流動的人,流動的DNA
智人約一萬年前開始定居種田以前,生活方式和尼安德塔人一樣,也習慣分為一小群一小群人活動,不長期定居在一個地點。有意思的是,舊石器時代已知少少的智人基因組,都不存在近親繁殖。
依賴採集、狩獵的生產方式下,每一群的人數都不多,近親配對好像很難避免。不過移動性高的人群,應該也常有機會互相交換人口,增加配對選項。從古代 DNA 看來,這是古早智人的普遍行為。
現有證據似乎告訴我們,遠比文明誕生更早以前,智人已經習慣刻意和血親以外的對象配對,或許可稱之為智人的「天性」,但是不清楚能追溯到多早。
智人如今僅有尼安德塔人一種比較對象,而尼安德塔人好像不排斥近親繁殖。有可能兩者的共同祖先已經會避免近親配對,尼安德塔人卻不再在意;也有可能這是智人較新的性擇模式,與尼安德塔人分家以後的某個時候才形成。

這也可以澄清一個疑惑。有個說法是,原始人只知道媽媽,不知道爸爸,因為小孩明確由媽媽生出,爸爸的功能卻不直接。根據古代 DNA 的證據判斷,此說很顯然錯誤。
如果隨機配對,一群人中勢必會有一定比例的人,父母為血緣近親。由結果反推,倘若都沒有的話,表示這群人都會刻意避免近親配對。
假如多數人都不知道爸爸是誰,實在難以想像要怎麼如此徹底的避免近親繁殖。反過來則合理得多:每個人都知道自己的爸爸媽媽是誰,擇偶時才能避開。
定居的人,設法讓 DNA 流動
一萬多年前開始,世界許多地方陸續有人定居下來,改為依靠種田營生。從流動性高的採集狩獵小群體,變成長期住在一處的小農村,人類的生活方式改變很大,這會影響配對習慣嗎?
人人採集狩獵的時期,每一群的人數都不多,但是習慣跑來跑去,有不少機會交換人口。新石器時代定居下來以後,初期的人口還是不多,卻失去流動性,只能從住在附近的有限對象中擇偶。如此一來,近親配對的機率應該會提高?
目前對此問題的探討不多。資訊比較多的案例,來自安那托利亞(現今的土耳其)一萬多年前,人口頂多數百的小農村遺址 Boncuklu、Pınarbaşı。這兒新石器時代初期的居民,多數在本地長大;可是遺傳上看來,都會避免近親繁殖。

具體狀況不明,本地與否是透過「鍶」的穩定同位素判斷,涵蓋的地理範圍不算太小。幾十公里遠的隔壁村,只要鍶同位素仍屬同一範圍,仍然會辨識為本地人。
不過我想這些線索應該足以支持,安那托利亞的人們邁入定居時代後,依然保持舊日的擇偶習慣,在有限的選項中盡量避免血親。但是近親繁殖也出現了。肥沃月灣西側的 Ba’ja 遺址(現今的約旦),至少有 1 位居民的父母為近親。
要提醒各位讀者,不同地方邁入定居的年代與狀況都不一樣,有時候差異很大,不可一概而論。
從城市到文明
隨著人口增長加上工作分化,漸漸有大型聚落誕生,有些或許可稱之為城市。人類發展可謂來到另一階段。
例如前述 Boncuklu、Pınarbaşı 遺址附近,就形成知名的加泰土丘(Çatalhöyük),數千年來都有數千人口居住。由鍶穩定同位素判斷,這兒多數人是土生土長,也有少量外來移民。
加泰土丘和我們習慣的「城市」有不少差異,卻昭示人類進入大量人口群聚的階段,各地一座又一座城市興起又衰落。長期保持數千人口的城市生活圈中,即使一輩子不出遠門,似乎也不難找到近親以外的異性配對。

當然在現代以前,世界各地的大部分人類並不住在人擠人的城市,而是人口密度更低的郊區與鄉村。不過倘若有心避免近親配對,應該不難達成。
目前為止重現於世的古代基因組,不論何時何地,大部分不是近親繁殖的產物。某文化的眾多樣本中,有時候能見到零星幾位,甚至是兄弟姊妹或親子間的極近親,但是都不普遍。
人口有限的海島,近親繁殖好像更容易發生。義大利南方的馬爾他島,在新石器時代確實如此;但是不列顛北部的奧克尼島,青銅時代僅管人口很少,依然能幾乎避免。
是人性的扭曲,還是財富的累積?
至今所知近親繁殖最常見的古代社會,是青銅時代的愛琴世界,也就是希臘及其外島,距今 3000 到 5000 多年前,愛琴海一帶的米諾斯等文化。薩拉米斯島(Salamis)等小島的比例較高,希臘大陸相對低,整體比例約 30% 之高。
- 延伸閱讀:愛琴世界,習慣親上加親?
取樣一定有偏差,真正的近親比例不好說,但是大概足以判斷青銅時代的愛琴世界,堂表兄弟姊妹等級的近親婚配習以為常,不只少量統治家族,而是全民普及的現象。

有史以來智人都會避免近親繁殖,為什麼愛琴人改變婚配方式?目前沒有答案。考古學家提出一個可能,種植橄欖之類的經濟作物,最好不要分割土地,而近親配對有助於保留土地,讓產業留在大家族內傳承。這聽起來合理,可惜缺乏更直接的證據。
社會中有人累積土地等資產,是人類發展的趨勢之一,而不論王公貴族或小地主,時常都有集中資產的需求。目前缺乏古代基因組的其他文化,是否也會見到類似愛琴世界的現象?我猜頗有可能,應該是有趣的探索方向。
隨著不同時空的樣本累積,加上容易操作的父母親緣分析軟體,未來「父母是否為近親」也許能成為古代基因組的標準化分析步驟,讓我們更方便認識人類的性擇。
延伸閱讀
- 羅馬尼亞3.5萬年前PM小姐,保有智人遺傳多樣性
- 舊石器時代採集狩獵小族群,懂得避免近親配對?
- 安那托利亞新石器時代早期,一起長眠的是親戚嗎?
- 克羅埃西亞史前遺傳史,大家都避免近親繁殖
- 愛爾蘭新石器時代,近親繁殖的統治階級?
- 歐洲邊緣的古代DNA,青銅時代超大量外來女生來到奧克尼島?
- 小河公主等4000年前新疆人,遺傳不是西方人,也不是現在的東方人
- 加勒比海一家親遺傳史
- 匈牙利外來政權,東方移民建立的阿瓦爾帝國
參考資料
- Scott, E. M., Halees, A., Itan, Y., Spencer, E. G., He, Y., Azab, M. A., … & Gleeson, J. G. (2016). Characterization of Greater Middle Eastern genetic variation for enhanced disease gene discovery. Nature genetics, 48(9), 1071-1076.
- Genomic landscape of the Greater Middle East
- Skov, L., Peyrégne, S., Popli, D., Iasi, L. N., Devièse, T., Slon, V., … & Peter, B. M. (2022). Genetic insights into the social organization of Neanderthals. Nature, 610(7932), 519-525.
- Sikora, M., Seguin-Orlando, A., Sousa, V. C., Albrechtsen, A., Korneliussen, T., Ko, A., … & Willerslev, E. (2017). Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers. Science, 358(6363), 659-662.
- Svensson, E., Günther, T., Hoischen, A., Hervella, M., Munters, A. R., Ioana, M., … & Jakobsson, M. (2021). Genome of Peştera Muierii skull shows high diversity and low mutational load in pre-glacial Europe. Current Biology, 31(14), 2973-2983.
- Pearson, J., Evans, J., Lamb, A., Baird, D., Hodder, I., Marciniak, A., … & Fernández-Domínguez, E. (2023). Mobility and kinship in the world’s first village societies. Proceedings of the National Academy of Sciences, 120(4), e2209480119.
- Yaka, R., Mapelli, I., Kaptan, D., Doğu, A., Chyleński, M., Erdal, Ö. D., … & Somel, M. (2021). Variable kinship patterns in Neolithic Anatolia revealed by ancient genomes. Current Biology, 31(11), 2455-2468.
- Wang, X., Skourtanioti, E., Benz, M., Gresky, J., Ilgner, J., Lucas, M., … & Stockhammer, P. W. (2023). Isotopic and DNA analyses reveal multiscale PPNB mobility and migration across Southeastern Anatolia and the Southern Levant. Proceedings of the National Academy of Sciences, 120(4), e2210611120.
- Cassidy, L. M., Maoldúin, R. Ó., Kador, T., Lynch, A., Jones, C., Woodman, P. C., … & Bradley, D. G. (2020). A dynastic elite in monumental Neolithic society. Nature, 582(7812), 384-388.
- Fowler, C., Olalde, I., Cummings, V., Armit, I., Büster, L., Cuthbert, S., … & Reich, D. (2022). A high-resolution picture of kinship practices in an Early Neolithic tomb. Nature, 601(7894), 584-587.
- Rivollat, M., Thomas, A., Ghesquière, E., Rohrlach, A. B., Späth, E., Pemonge, M. H., … & Deguilloux, M. F. (2022). Ancient DNA gives new insights into a Norman Neolithic monumental cemetery dedicated to male elites. Proceedings of the National Academy of Sciences, 119(18), e2120786119.
- Dulias, K., Foody, M. G. B., Justeau, P., Silva, M., Martiniano, R., Oteo-García, G., … & Richards, M. B. (2022). Ancient DNA at the edge of the world: Continental immigration and the persistence of Neolithic male lineages in Bronze Age Orkney. Proceedings of the National Academy of Sciences, 119(8), e2108001119.
- Ariano, B., Mattiangeli, V., Breslin, E. M., Parkinson, E. W., McLaughlin, T. R., Thompson, J. E., … & Bradley, D. G. (2022). Ancient Maltese genomes and the genetic geography of Neolithic Europe. Current Biology, 32(12), 2668-2680.
- Freilich, S., Ringbauer, H., Los, D., Novak, M., Pavičić, D. T., Schiffels, S., & Pinhasi, R. (2021). Reconstructing genetic histories and social organisation in Neolithic and Bronze Age Croatia. Scientific Reports, 11(1), 16729.
- Gnecchi-Ruscone, G. A., Szecsenyi-Nagy, A., Koncz, I., Csiky, G., Racz, Z., Rohrlach, A. B., … & Krause, J. (2022). Ancient genomes reveal origin and rapid trans-Eurasian migration of 7th century Avar elites. Cell, 185(8), 1402-1413.
- Fernandes, D. M., Sirak, K. A., Ringbauer, H., Sedig, J., Rohland, N., Cheronet, O., … & Reich, D. (2021). A genetic history of the pre-contact Caribbean. Nature, 590(7844), 103-110.
- Zhang, F., Ning, C., Scott, A., Fu, Q., Bjørn, R., Li, W., … & Cui, Y. (2021). The genomic origins of the Bronze Age Tarim Basin mummies. Nature, 599(7884), 256-261.
- Skourtanioti, E., Ringbauer, H., Gnecchi Ruscone, G. A., Bianco, R. A., Burri, M., Freund, C., … & Stockhammer, P. W. (2023). Ancient DNA reveals admixture history and endogamy in the prehistoric Aegean. Nature Ecology & Evolution, 1-14.
本文亦刊載於作者部落格《盲眼的尼安德塔石匠》暨其 facebook 同名專頁。