- 文/黃湘芹、謝若微、李映漾、陳柏仰|中央研究院植物暨微生物學研究所

可以在不改變 DNA 的狀況下,調整性狀?——表觀遺傳調控,幫助植物快速適應環境變化
DNA 是生物細胞內攜帶遺傳訊息的物質,當 DNA 發生變異時,會影響基因的表現進而改變性狀。但很多生物也可以在不改變 DNA 的情況下調節基因表現影響性狀,此方式稱為表觀遺傳調控,其中常見的機制包括 DNA 甲基化、組蛋白修飾、小分子 RNA 等。
其中「DNA 甲基化」為在 DNA 特定位置上添加甲基的化學修飾,當基因前端的區域——啟動子被高度甲基化時,常會導致基因表現量較低。
而「組蛋白修飾」是針對被 DNA 纏繞的蛋白質——組蛋白,在其尾端上做的各種修飾,如乙醯化、甲基化、磷酸化等,這些修飾會影響 DNA 纏繞的緊密程度,進而加強或抑制基因表現[1]。另外,由長度約為 18 到 30 個核苷酸構成的「小分子 RNA」,也會抑制基因表現。
對生物而言,表觀遺傳調控提供生物在基因序列突變外,另一種有效適應環境變化的反應方法。而這樣的反應對植物特別重要,它能幫助植物在面對氣候、環境快速變化時,迅速調整基因表現讓植物得以生存。
如果將表觀遺傳運用在改良作物性狀上,由於不需外來基因插入或是基因編輯,便能達到基因表現的變化,因此大幅減少食物安全上的諸多考量,免除基改作物對人體健康疑慮的爭議性,在農業發展上相對有利。
目前在作物中已有不少研究,分析基因體上特定位置的表觀遺傳變異,與抵抗逆境性狀之間的關聯性;例如在稻米基因體上,已發現數個特定位置的 DNA 甲基化程度與抗旱[2]、抗缺鐵[3]甚至碳儲存有顯著的關聯性。

在番茄裡也發現,由小分子 RNA 對特定位點的基因調控,可影響番茄外型及抗旱性狀。顯示透過影響表觀遺傳機制,的確有機會用來培育出具有優良性狀的作物。
如何運用在作物改良上?
當應用於作物改良時,偵測表觀遺傳變異與性狀之間的關係為首要任務,其中一種用來偵測表觀遺傳變異的策略仰賴的是近年才逐漸普遍化的「全基因體定序」。由於每個作物的基因體序列不同,需逐一檢視不同作物在各種逆境條件下產生的表觀遺傳變異,然而在技術與基因體資料分析上仍是挑戰。現階段而言,利用表觀遺傳進行作物改良,雖有潛力但未能普及[4]。
利用全基因體定序偵測表觀遺傳變異(圖表一):先透過外在刺激誘導表觀基因座產生變異,接著藉由分析眾多植株間表觀基因座變異的差別,並計算其與目標性狀的關聯性,進而推定能產生目標性狀的表觀基因座。
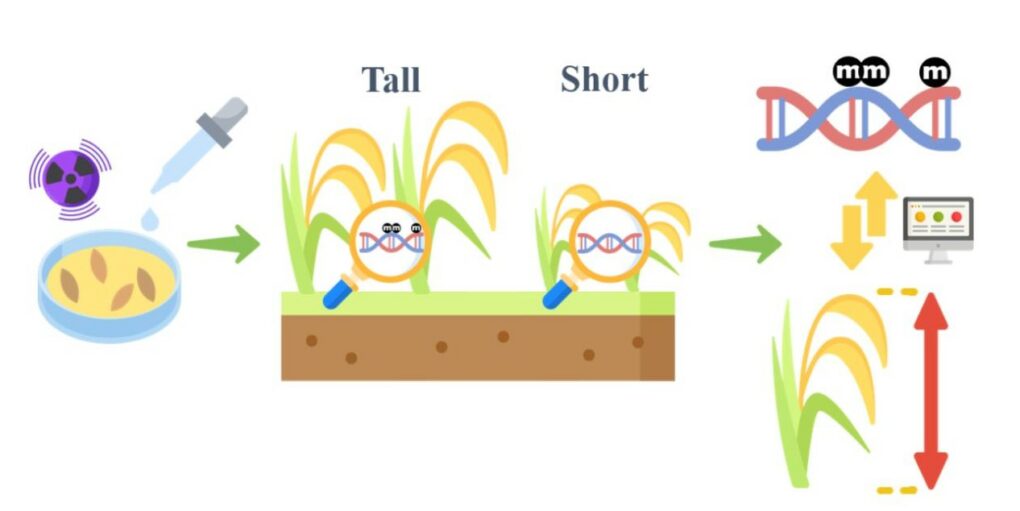
在已知可誘導表觀基因座的策略中,以 DNA 甲基化為例 ,透過伽馬射線照射、DNA 甲基轉移酶抑制劑以及組織培養,皆可在稻米基因體產生隨機且有效的 DNA 甲基化變化[4]。
種子如果曝露於伽馬放射線環境下,或是浸泡在含有 DNA 甲基轉移酶抑制劑的水溶液中,均會造成基因體去甲基化,而去甲基化的程度會隨著放射線強度或是 DNA 甲基轉移酶抑制劑的添加量而不同;如果同時使用上述兩種方法處理種子,則會對於去甲基化有加乘效果[5]。

除了水稻以外,玉米基因體上特定位點的表觀遺傳因子變化,可改變其對於熱逆境的耐受性。玉米在幼苗時期,如果受到短暫熱處理,便能促進與基因表現有關的組蛋白修飾,使得葉片的葉綠素含量與活性氧物質提高,以增強玉米在高溫環境下的耐受性[5]。
面臨的挑戰——表觀遺傳變異重現與否
表觀遺傳變異與基因變異主要的不同在於其不穩定性,由於細胞有自我修復機制,因此表觀遺傳變異在細胞複製前、後未必能維持;此外,世代遺傳間的「表觀遺傳重組」(epigenetic reprogramming)會重置表觀遺傳的分佈,使得親代的變異未必能完整保留到子代。
儘管如此,不少研究仍發現部分表觀遺傳變異可以被遺傳至下一代。以茄科中常用的嫁接作物番茄、茄子與辣椒為例,這類的種間嫁接會影響 DNA 甲基轉移酶表現量,進而大規模影響接穗中的 DNA 甲基化分佈,其中有部分 DNA 甲基化的變動被證實可維持至下一代[5]。
綜合上述,應用表觀遺傳在作物改良上需特別確認變異在跨世代間的一致性;植株進行處理後所產生的表觀遺傳變異,是否能在性狀植株或甚至下一代重現,以確保有效的作物改良。
甜椒的跳躍基因與 DNA 甲基化

甜椒 (Capsicum species)的基因體解序後,發現當中的跳躍基因(可以在基因體上移動的 DNA 序列)不僅增加了甜椒的多樣性,也能決定轉錄活性高的真染色質及不具轉錄活性的異染色質在基因體上的分佈[6],從而廣泛影響基因調控。
已知 DNA 甲基化是控制跳躍基因的主要因子,已有研究指出,甜椒基因上 DNA 甲基化程度的增加,與發芽、果實成熟及抗鹽性狀都有顯著相關[7][8];顯示透過刺激產生的 DNA 甲基化重新分佈,極可能影響跳躍基因的活性,進而引導出優良性狀。
目前甜椒的基因體資料已完備,其重要性狀與表觀遺傳變化密切相關,被視為可積極利用表觀遺傳進行改良的高經濟價值作物之一。
翻開作物育種的新篇章
綜合以上,分析及尋找與目標性狀相關的表觀基因座並不容易,需要結合農藝學、基因體學及生物資訊學的知識與技術,考量表觀遺傳變異的不穩定性因素,為實現可代代相傳的作物改良,需要了解不同植物的基因體中有哪些特定的表觀遺傳變異能夠穩定傳到下一代。因此,若要使用表觀遺傳改良作物,雖有理想但非一蹴可及。
目前主流的基因改造工程,在食品、環境、與生物安全上有著錯綜複雜的影響,僅透過調控基因表現以達到性狀改良的表觀遺傳,更能消除大眾對於作物改良的疑慮。現今對於表觀遺傳的研究資料已經越來越多,在植物面臨逆境時,表觀遺傳能有效且迅速地幫助植物適應環境。在未來環境更加極端的情況下,生產作物將會面臨更嚴峻的挑戰,如何繼續維持高產量,成為農民及研究者必須解決的問題之一。
表觀遺傳調控提供植物學家與農民新的作物改良方法,儘管當前的流程尚不完善,也有許多困難需一一克服,但看好其在未來為作物育種開啟新篇章。
參考資料
- Tirnaz, S. & Batley, J. (2019). Epigenetics: potentials and challenges in crop breeding. Molecular Plant, 18, 1309–1311.
- Sapna, H., Ashwini, N., Ramesh, S. & Nataraja, K. N. (2020). Assessment of DNA methylation pattern under drought stress using methylation-sensitive randomly amplified polymorphism analysis in rice. Plant Genetic Resour Charact Util, 18, 222–230.
- Sun, S., Zhu, J., Guo, R., Whelan, J. & Shou, H. (2021). DNA methylation is involved in acclimation to iron deficiency in rice (Oryza sativa). Plant J, doi:10.1111/tpj.15318.
- Springer, N. M. & Schmitz, R. J. (2017). Exploiting induced and natural epigenetic variation for crop improvement. Nat Rev Genet, 18, 563–575.
- Varotto, S. et al. (2020). Epigenetics: possible applications in climate-smart crop breeding. J Exp Bot, 71, 5223–5236.
- Kim, S. et al. (2014). Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat Genet, 46, 270–278.
- Xiao, K. et al. (2020). DNA methylation is involved in the regulation of pepper fruit ripening and interacts with phytohormones. J Exp Bot, 71, 1928–1942.
- Portis, E., Acquadro, A., Comino, C. & Lanteri, S. (2004). Analysis of DNA methylation during germination of pepper (Capsicum annuum L.) seeds using methylation-sensitive amplification polymorphism (MSAP). Plant Sci, 166, 169–178.