由化石、遺骸等材料獲取古代 DNA,是探索生物遺傳史的利器。2022 年一篇論文報告大量古代狼的基因組,探討狼群的變遷。
狗源自於狼,對古代狼的研究,是否也能釐清狗在哪兒馴化?盡管這項研究沒有提供直接的明確答案,依然帶來有用的線索。

未滿 2.3 萬年的狼,血緣主要源自西伯利亞
中文稱之為「狼」的動物有好幾款,狗的祖先是灰狼(grey wolf,學名 Canis lupus),主要住在北美洲,以及歐亞大陸靠北邊的區域,歐洲、中東、中亞、北亞、東亞。這項研究獲得 66 個新的古代基因組,加上之前發表過一共 72 個,覆蓋率介於 0.02 到 13。
最古老的樣本距今約 10 萬年,大部分地點位於歐洲、西伯利亞的東北部、北美洲。獲得每一個地區,不同年代的大批樣本,便能比較狼在不同時間、空間的血緣變化。
不同地區一直都有狼,假如各地的狼不太交流,那麼每一個地區的狼,遺傳上都會更接近同一地區,更早與更晚的狼,和其他地區的同類差異較大。然而比對得知,狼的血緣主要取決於時代,而非地點。
比方說歐洲 1 萬年前的狼,和 1 萬年前的美洲同類比較近,卻和 3 萬年前的歐洲狼比較疏遠。由此推論,古代各地狼群間的遺傳流動應該非常頻繁,沒有某地狼群孤立太久。
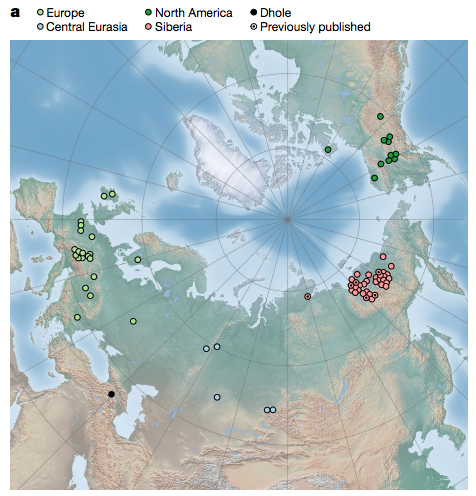
最明確的案例發生在距今 2.3 到 2.8 萬年前。比 2.3 萬年更晚的狼,和比 2.8 萬年更早的狼,遺傳上各自形成較近的一群。這是由於源自西伯利亞的血緣,向其他地區單向輸出所致。
晚於 2.3 萬年,各地的狼有很大比例血緣,能追溯到 2 萬多年前的西伯利亞狼群。歐洲古代狼群仍保留 10 到 40% 更早的血緣,沒有被完全取代。等到最近 1 萬年內,歐洲狼群的 DNA 又往西伯利亞、中國流動。
北美洲換過新血,再度與郊狼混血
相比之下,北美洲早於 2.3 萬年的血緣完全消失不見,徹底換上一批新血,和當地更早的同類可以說是不同的遺傳族群,光憑化石根本無從得知。
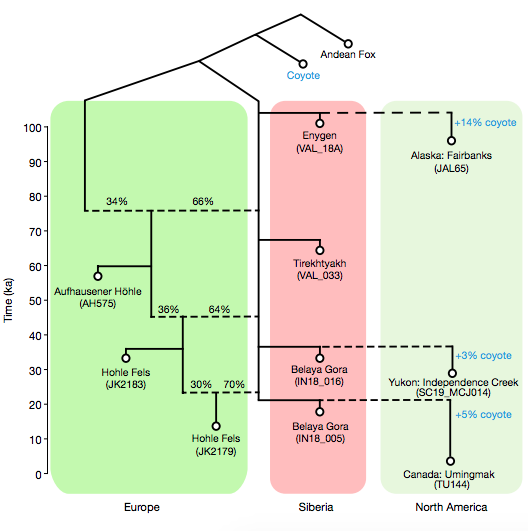
北美洲另有一個犬科物種:郊狼(coyote,學名 Canis latrans),和灰狼可以生產後代。兩者遺傳上約在 70 萬年前分家,至少 10 萬年前便陸續有遺傳交流。
北美洲晚於 2.3 萬年的狼,血緣皆能追溯到西伯利亞近期的移民;如今北美洲的狼群,可以視為前述血緣加上 10 到 20% 郊狼的合體。換句話說,北美洲比較早的狼就有郊狼血緣,全滅換過一批以後,很快又與郊狼混血。
北美洲的狼皆配備郊狼成分,而歐亞大陸所有的狼都缺乏,可見狼群向美洲的遷徙是單行道,只有從亞洲向美洲移民,沒有再回來的。
狼遺傳適應的存在感迅速躍升,比狗狗馴化更早
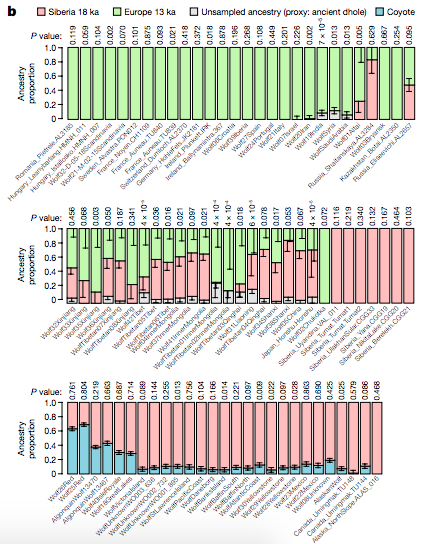
生物的 DNA 不斷改變,和外界環境互動之下,有些遺傳變異顯得有利,存在感上升。根據論文的分析辦法,在最近 10 萬年狼的基因組上偵測到 24 處遺傳適應。
最強烈的訊號位於第 25 號染色體的 IFT88 基因附近,距今 3 到 4 萬年前間,從 0% 直接躍升為 100%。此一基因和頭骨型態有關,但是不清楚對狼與狗的具體作用。其下游 2.5 Mb 處還偵測到另一個強烈訊號,2 到 4 萬年前間躍升為 100%。
除此之外還有幾處 DNA 變異,於 2 到 4 萬多年前存在感明顯增加。據此判斷在那個時期,各地的狼群有不少遺傳交流。而這些可能有利於狼適應的遺傳變異,狗狗也有配備,推測這些情慾流動的時刻,或許早於狗狗馴化的時間點。
所以狗到底什麼時候馴化的?多年下來也沒有一個很明確的答案,加上新研究的證據還是沒有,但是真相或許已經呼之欲出。
狗狗馴化真的是超級難題
狗的馴化是個超級難題,不是缺乏證據,而是比起其他馴化生物,狗明明有一大堆證據,卻互相矛盾,無法更加釐清問題。

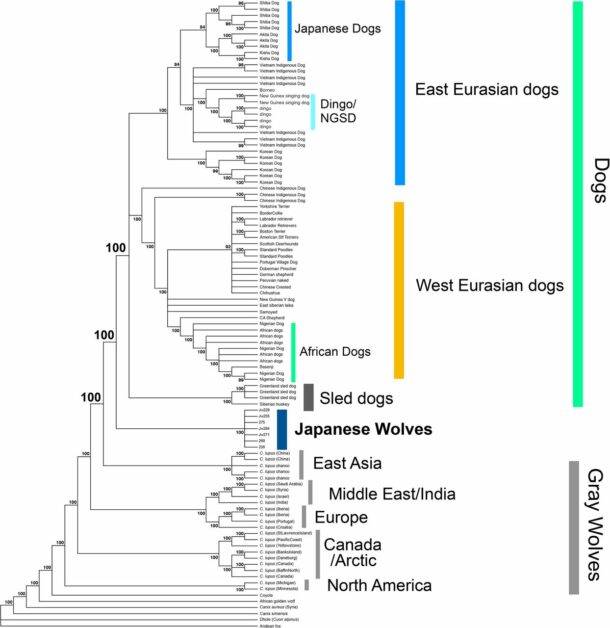
每一種馴化生物,都有野生的近親。野生近親中的一群後來衍生出馴化生物,因此馴化生物的直系祖先那一群,在演化樹上會較為接近馴化生物。例如野生的斑貓(Felis silvestris)有 5 個亞種,其中的非洲野貓(Felis silvestris lybica)衍生出馴化貓。
狗的狀況完全不一樣。將狗與狼擺在一塊畫演化樹,所有的狗自成一群,各地狼群也被歸類為另一群,兩群平行。過往通常解釋為:馴化為狗的那群狼已經滅團,所以我們見不到和狗在同一群的狼。
然而,這回加入大批不同時間、空間的古狼以後,狗的直系祖先狼依然不見蹤影。最接近狗狗的是距今 1.3 到 2.3 萬年前的西伯利亞古狼(也就是隨後各地所有狼的祖先),可是牠們們依然不是狗的直系祖先,是平行關系。
由此推敲,狗狗的直系祖先狼,和西伯利亞古狼在遺傳上應該早於 2.3 萬年前分家,否則演化樹上,狗就會在一群狼的內部。但是應該沒有早太多,因為當時兩者的差異還很有限,比其他地區的狼更小。
最初的狗於「東方」馴化?
仔細比較,狗的血緣更接近如今地理上偏歐亞大陸東方的狼,論文藉此推論,狗的馴化應該發生在「東方」,但是具體位置不明。
如今所有的狗,都缺乏早於 2.3 萬年歐洲狼的成分,歐洲為起源地的可能性,幾乎可以排除。而晚於 2.3 萬年的歐洲狼,依然小部分繼承前輩血緣,是牠們與狗差異較多的原因之一。
兩萬多年前發生什麼事呢?距今 1.9 到 2.6 萬年左右,全球進入酷寒的冰河時期,稱為末次冰盛期(Last Glacial Maximum,縮寫 LGM),大幅限縮生物的發展空間。對照狼的演化史,在此之後各地族群都被西伯利亞的狼群取代。
末次冰盛期之際,各地狼群很可能被切割開來,缺少遺傳交流機會,各自損失慘重,例如北美洲就全面滅團。身為狗狗直系祖先那群狼,或許當時也被孤立,更有機會與人類發生關係,造就馴化狗的契機。受到人擇之後,這支血脈與其他的狼在遺傳上明顯分開。
如果地點不是西伯利亞,大概也在不遠處,我猜是西伯利亞南部、華北、蒙古到中亞一帶。回答狗狗起源這個難題,這兒 2 到 3 萬多年前的化石,或許就保存著夢寐以求的基因組。
狗有兩地狼的血緣,但馴化是一次或兩次?
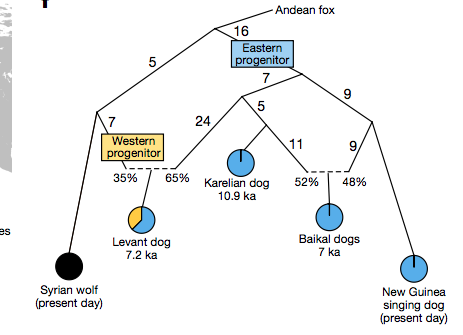
另一件有意思的發現是,除了上述血脈,狗狗們還具有另一款不同的血緣,遺傳上最接近現代中東到南亞一帶的狼群,姑且稱之為「西狗血緣」。
用敘利亞現代狼作代表,估計 7200 年前中東同一地區的古狗,配備 56% 類似的血緣。這個數字誤差不小,看看就好,但是足以肯定西狗血緣至少在 7200 年前已經存在。
相較於前述與西伯利亞古狼關係密切的「東狗血緣」,「西狗血緣」來自另一群古狼,牠們不住在歐洲,可能位於中東到南亞一帶,大部份血緣應該也源自 2 萬年前的西伯利亞古狼,只是分家年代晚於東狗血緣。
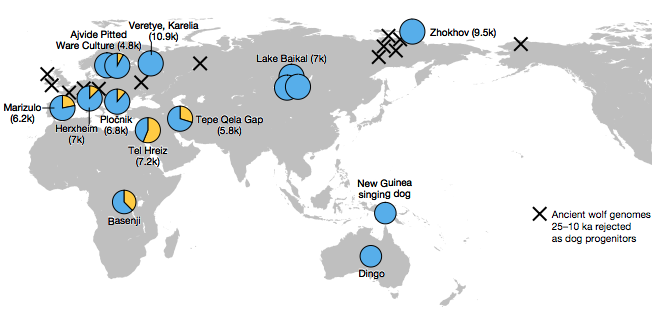
由此推敲,有批狼在東方變成狗以後,西方或許又發生過一次獨立的馴化,可是也有機會是東方狗到達以後,與當地狼大幅合體。
一個論點是:狗在東邊馴化一次,後來又融入西邊的狼。另一個論點是:狗在東邊、西邊各馴化一次。兩者皆符合目前的證據,隨著後續的 DNA 流動,兩款祖源都成為如今多數狗狗的一部分。
遠離歐亞大陸的新幾內亞唱犬(New Guinea singing dog)、澳洲野犬(dingo),都缺乏西狗血緣;牠們的祖先超過一萬年前便形成獨立遺傳支系,後來某個時刻又渡海抵達新幾內亞、澳洲。
考量這件事,我猜狗只在距今 2.6 萬年以前與過後的幾千年期間,於歐亞大陸偏東邊明確馴化一次,後來再傳播到各地;傳向東南方,新幾內亞唱犬的祖先一直獨立發展,缺乏西狗血緣;傳到歐亞大陸西邊,一萬年內的狗則與當地狼群合體,融入大量西狗血緣。不過目前這只是公堂上的假設。
狗狗的起源與演變,仍需要更多證據才能明確解答。不過這項研究的主角其實是狼,光是這方面獲得的新知便很值得學習。
延伸閱讀
參考資料
- Bergström, A., Stanton, D. W., Taron, U. H., Frantz, L., Sinding, M. H. S., Ersmark, E., … & Skoglund, P. (2022). Grey wolf genomic history reveals a dual ancestry of dogs Nature, 1-8.
- Ice Age wolf DNA reveals dogs trace ancestry to two separate wolf populations
- Ancient wolves give clues to origins of dogs
本文亦刊載於作者部落格《盲眼的尼安德塔石匠》暨其 facebook 同名專頁。