


是的,寄生獸電影的完結篇都要上映了,關於寄生獸的寄生生物學也該告個段落收尾了。
之前兩篇文章,談到的分別是寄生獸的生活史跟宿主以及感染生態學兩面向。這一次,我們從寄生蟲跟宿主互動的角度,來仔細瞧瞧寄生獸感染宿主之後在宿主體內發生的事情。
(本篇可惜還是不由講談社或東立出版社贊助)
如同第一篇寄生獸的文章所提到的,寄生蟲通常是有宿主專一性的,就算不是專一到屬或種,多半也是專一在某個類群。更進一步的說,寄生蟲既然要奪取宿主的資源,大張旗鼓的跟寄主硬碰硬恐怕只會弄得玉石俱焚,多半討不了好。所以,寄生蟲們相對於宿主,絕大多數都是體型上小得多,這樣才方便悄悄的待在宿主身上行五鬼搬運蠶食鯨吞之法。而既然不是像鬼魂附身一樣侵佔整個宿主身體,那麼寄生蟲要待在宿主身上的什麼部位其實也是有所選擇的。
好比說跳蚤這樣的體外寄生蟲好了,其宿主的尺寸如牛羊家畜與跳蚤相比何止九牛與一毛,但是跳蚤在宿主身上並不是隨處都好,而是有著明顯的身體部位偏好。這樣的偏好有可能是取決於跳蚤自身(例如該處的宿主皮膚較薄、毛髮較濃密、微血管分佈較淺等),或者是受到宿主抓搔理毛行為干擾的結果(例如該身體部位比較宿主理毛理不到)。

體外寄生蟲都有身體部位的偏好了,體內寄生蟲當然也如是,同樣有宿主部位專一性。又既然宿主相對寄生蟲來說這麼龐大,要從寄生蟲進入的位置移動到目的地,總是要花上一番功夫。簡單一點的做法是利用宿主本身的循環系統搭順風車到目的地去,辛苦一點則是寄生蟲得自己主動找路找孔走現成管道過去,或甚至得要披荊斬棘在宿主的組織裡一路殺出一條通道。
然而,既然我們現在有了GPS都還是可能會迷路,無論是搭順風車、主動找路或是自己開路,在宿主體內移行的寄生蟲當然也難免迷途或受到阻礙。不過,萬一迷了途走錯路,寄生蟲大概就很難有機會回到正確的目的地,又受限於本身的發育進程,走錯路的寄生蟲大概就只能在錯誤的宿主部位徒呼負負,就算僥倖沒有被宿主的免疫系統攻擊而夭折,大概也就只能發育停滯空餘恨或是被宿主排除了。
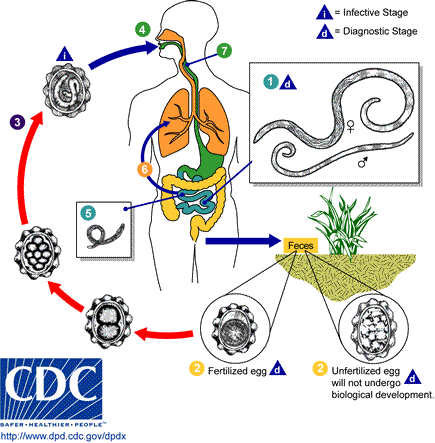
以寄生在消化道裡頭的蛔蟲為例,成蟲蛔蟲絕大多數住在空腸段,排出來的卵跟著宿主糞便到體外,卵中的胚胎發育成具有感染力的二齡幼蟲如果又有幸被宿主吃下肚裡,則在小腸段孵化並穿透腸壁,順著宿主的循環系統搭順風車移動到肺部,然後穿出血管住在肺泡裡。等到幼蟲發育成熟後,就會主動的爬出肺部上到氣管,然後在咽喉處引起一陣輕咳,再順勢被吞回消化道,經過胃部回到小腸。
在這個生活史的旅途中,搭順風車的蛔蟲幼蟲有可能下錯站,該在肺部下車卻到了腎臟或腦部下車,於是也就只能在不適合的宿主部位等死,並且在死前把宿主的身體搞得一團糟,視下錯車的部位導致腎衰竭、腦炎、癲癇或製造一些嚴重免疫反應之類。相較之下,主動移動的成熟蛔蟲跑錯路線,頂多也只是從宿主的口鼻部被打噴嚏或咳出來而已,倒是對宿主沒有太多傷害,只是製造很噁的回憶而已。
回頭來看寄生獸,雖然如上一篇寄生獸文章所觀察描述的,寄生獸是由現成孔洞(例如耳朵)進入宿主體內,而且一路向上的進入宿主腦袋然後控制宿主。也就是說,寄生獸的幼蟲除了在現成管道裡主動移動之外,也得要有一段披荊斬棘分筋錯骨的路程,才能進到目的地腦袋裡。雖然說口鼻耳孔進入管道之後距離腦袋很近,但腦袋畢竟還是有頭骨密密保護著的,而且你看這寄生獸幼蟲這麼大一條,就算它們超級伸縮自在,穿過頭骨的時候應該還是「很有感覺」。畢竟,小小的一般寄生蟲在皮下造成的幼蟲移行症(larva migrans)就已經很不舒服了,更何況是那麼大隻的寄生獸幼蟲穿過腦殼呢是不是。

不過,就跟一般寄生蟲一樣,寄生獸入侵宿主的過程也不一定是都那麼順利的。就好像寄生獸的主角米奇一樣,如果沒辦法從口鼻耳孔進入宿主,寄生獸也是可以硬著頭皮從其他地方入侵,然後一路走皮下向頭部移動。一般來說,寄生蟲通常沒有這麼大的本事能夠轉換入侵路徑,但大概是因為寄生獸的入侵方式結合了自然孔道入侵和組織間移行吧,所以當沒有自然孔道可走的時候也就直接走組織間移行的路線了。而這沿著皮下移行的景象,除了類似於剛剛提過的皮下幼蟲移行症之外,其實更像是麥地那龍線蟲(Dracunulus medinensis)在皮膚底下移行的感覺…

(想知道麥地那龍線蟲並且不怕蟲蟲上身的噁心感請參閱維基百科,生活史示意,還有播放下面的影片)

總之,走錯路的寄生蟲一般來說就是死路一條,但這寄生獸顯然比較特別一點。雖然說寄生獸主要侵佔的部位是腦部(或該說是整個頭部?),但是萬一不得已,也是可以屈居其他宿主部位的,好比說主角新一的右手,或是另一個人類宿主的下巴頸胸部位。這一點,寄生獸就比現實生活中的寄生蟲彈性得多。
而且屈居非目標部位的理由,居然還可以是因為宿主即將死去,所以先與宿主融合以拯救宿主?是啦,寄生蟲跟宿主其實是命運共同體,真的要長治久安的話,寄生蟲的確是該讓宿主好好活著,這樣才能有源源不絕的資源可供剝削。但在自然界中,信手拈來也是有一狗票的寄生蟲,當它們為了完成生活史的時候就會操縱宿主去自殺(例如第一篇文章提到的弓漿蟲操縱老鼠、絛蟲操縱蝸牛、或是底下的兩個影片),完全不留情的啊。
說到底,蟲不為己天誅地滅,宿主不過是寄生蟲的資源罷了,一切還是以寄生蟲的福利為依歸,而像寄生獸這樣神通廣大考慮到未來又有能力拯救宿主於溺水的寄生蟲,還真的沒見過哪。但話說回來,寄生獸雖然能力強大,但要是遇上解決不了的宿主的傷害(好比說車禍重傷或宿主體內有毒素入侵)時,寄生獸還是會拋下宿主另覓生路的。這一點,寄生獸又跟寄生蟲沒啥兩樣。就好像,當一隻有鐵線蟲寄生體內的蟋蟀不幸被捕食者吃掉的時候,這鐵線蟲並不是「跟宿主一起共存亡」或什麼「寄宿同心其力斷金」,而是自顧性命能跑就跑。於是,出現「蛙口脫逃」或「魚鰓脫逃」這樣的場面,其實也就不難理解了。
最後,負責解剖和分析寄生獸的醫師由井是這樣描述的:
寄生獸的細胞本質跟人類非常相似,但是特點在於他們像是「會思考的肌肉」,於是能瞬間變成橡膠般柔軟、有彈性或鋼鐵般堅硬、鋒利。
這樣的敘述讓我想到黏菌,但黏菌並沒有那麼大的本事可以隨心所欲變化材質特性,頂多只有從隨意變形的變形體轉成有固定形狀的子實體罷。不過既然這變形體看來軟趴趴而子實體又要有固定形狀想來也是稍有硬度,或許黏菌的這個特質也是寄生獸作者取材的對象吧。
但重點是,黏菌不是寄生蟲啊,黏菌不是寄生蟲啊,黏菌不是寄生蟲啊啊啊啊啊。(為什麼要講三次大家應該都知道了)


文章寫到這裡,最後一個寄生獸的奧妙之處,其實是在於「宿主部位取代」這件事情。雖然漫畫裡說寄生獸主要是入侵腦部,但精確一點來看,其實應該是『取代宿主頭部』才對。不管是從寄生獸在宿主之間轉換時要砍掉新宿主的肢體部位,或是後來軍方要消滅寄生獸時以X光掃描可疑群眾的畫面,其實都明示了寄生獸根本不是「侵佔」宿主身體部位而已,而是整個把宿主的部位消化掉並且取而代之。
這一點,在寄生蟲案例上幾乎是少之又少,畢竟寄生蟲雖然住在宿主身上,但終究是涇渭分明的兩個生物個體,寄生蟲在寄生之後把自己身體簡單化或部份融入宿主身體部位的例子不是沒有(例如下圖的寄生橈腳類),但要越俎代庖的「變成」宿主身體部位的模樣還要執行正常功能(寄生獸變成的頭部是可以吃喝視聽嗅外加操控整個宿主身體的啊),那還真的是幾乎沒有。


是的,幾乎沒有,但還是有。生物的奧妙之處,就在於總是有各種古怪的案例可供作者取材。要說寄生蟲「取代」了宿主的身體部位而且還執行功能,那就一定要看縮頭魚虱這種「目前已知唯一完全取代宿主器官的寄生方式」的寄生等足目了。
縮頭魚虱(Cymothoa exigua)有一票的近親種,大家都是魚體上或口腔內的寄生蟲,但不知為何,就只有縮頭魚虱演化成這樣,不但進駐了宿主魚的口腔,巴住了宿主的舌頭吸血,讓宿主的舌頭萎縮掉,居然還佛心來著的就抓在原本舌頭的位置上,取代了舌頭的功能?!各位不覺得這有點像是小偷進了家門,搜刮財物之餘居然還幫你整理家務照顧房子一樣,搞到後來都不知道誰是屋主,還有該不該感謝對方了……
瞧瞧這縮頭魚虱那尖尖的爪子,詭異的取代宿主部位的習性,我真的覺得寄生獸應該有從中得到靈感吧?你說是嗎?
(寄生獸全系列文章完,謝謝收看,我們後會有期。)






























-200x200.jpg)




{kind=link}
{kind=link}