
- 文 / 李承叡|臺灣大學生態學與演化生物學研究所
你知道西洋芥末與東洋山葵的辣味源自同一類化合物嗎?硫代葡萄糖苷(glucosinolate)常見於十字花科植物(芥末、山葵、白菜、高麗菜、花椰菜、芥菜等),當植物細胞被破壞後,硫代葡萄糖苷會被分解,而其下游產物就是芥末及山葵刺激性味道的主要來源(圖一)。
就像許多植物次級代謝物一樣,這類化合物對植食性昆蟲具有毒性。因此,不像人類吃芥末是為了尋求刺激,植物體內芥末的含量與種類對昆蟲來說可是生與死的差別!

左圖/Wikipedia、右圖/Wikipedia
當然就如同自然界的大部分性狀,硫代葡萄糖苷不會只有一類。不同的硫代葡萄糖苷源自不同種類的氨基酸前驅物。在化學生態學(chemical ecology)與演化遺傳學(evolutionary genetics)的領域,科學家在意的是為什麼會有化合物多樣性,是什麼因素維持這些多樣性,背後又是發生了怎樣的遺傳變異?
基因重複導致化合物多樣性
在北美洲的野生植物 Boechera stricta,硫代葡萄糖苷的多樣性主要來自兩大類化合物比例上的差別:一類由甲硫胺酸(methionine)為前驅物(Met GS)、另一類由支鏈氨基酸(branched-chain amino acid)為前驅物所產生(BC GS)。
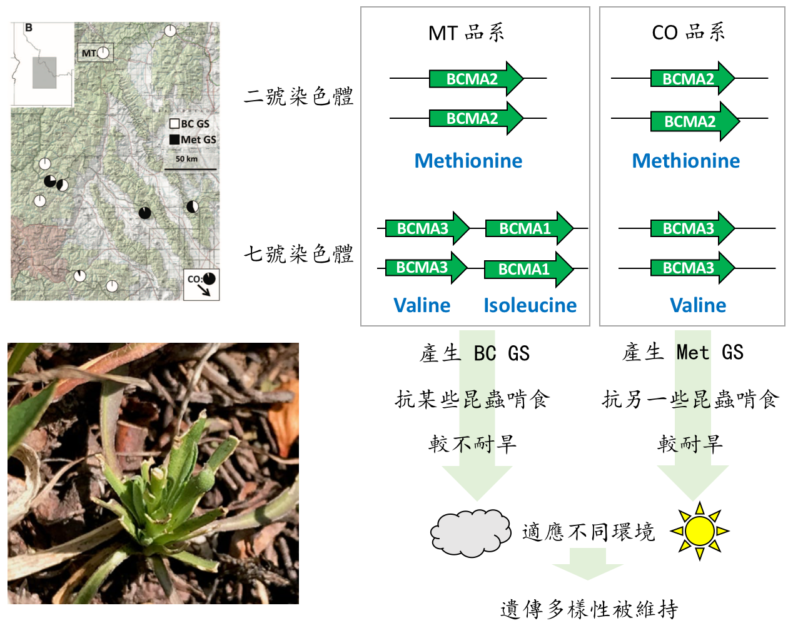
科學家們首先發現野外族群這兩類化合物的比例大不相同(圖二),並透過基因定位找出了控制這個差異的基因座 [1]。這個基因(BCMA)產出的酵素參與硫代葡萄糖苷生成的關鍵第一步:突變讓酵素有不同的受質偏好性,抓了不同的氨基酸送往後續生化反應,產生不同的硫代葡萄糖苷。
但和一般孟德爾遺傳的例子不同,這並不是單一基因和兩個對偶基因這麼簡單!在這邊,遺傳多樣性源自基因重複(gene duplication)。
基因重複與 DNA 複製(DNA replication)不同,基因重複會把基因體的一個小片段從一份複製成許多份,同時基因體其他部位的基因含量不變。
但重複後的這幾份基因功能不是多餘的嗎?因此我們常常可以發現同一個基因在產生重複之後,多餘的那些份數可能經過突變讓其完全失去功能,或與原本基因負責不同組織的表現,甚至可能產生全新的功能!
在人體內最有名的例子是球蛋白(globin)基因:它從遠古時的一個基因,經過大量重複及功能改變後產生肌紅蛋白(myoglobin)及血紅素(hemoglobin)的不同次單元基因,各自負責在不同組織和發育時期與氧結合。
回到野生芥末,這個 BCMA 基因也發生過許多次基因重複(圖二):在二號染色體的祖先型 BCMA2 偏好抓取甲硫胺酸。經過基因重複、跳躍到七號染色體、並發生突變之後,產生了也會抓取支鏈氨基酸的其他拷貝:BCMA1 使用異白胺酸(isoleucine),BCMA3 使用纈胺酸(valine)。
我們發現體內含有這三個 BCMA 基因(BCMA1、BCMA2、BCMA3)的品系 MT,生產出「由支鏈氨基酸衍生的硫代葡萄糖苷」(BC GS)的比例較高。
而在品系 CO,BCMA1 基因後來又被刪除了,因此 BC GS 比例極低[1]。應用分子演化學演算法,我們發現基因重複後受到強烈天擇壓力、演化快速的氨基酸位點正好位於此酵素辨認受質的區域。最後,我們的合作者利用生化分析,證實了只要把祖先型 BCMA2 上的這些氨基酸位點,置換為 BCMA1 或 BCMA3 上面的氨基酸,就足以增加其抓取支鏈氨基酸的機會[1]!因此,我們證明了基因重複、突變產生的新功能、及後續基因丟失,會增加硫代葡萄糖苷的多樣性。
但是,這樣的突變對生態和演化上有什麼意義呢?
基因重複影響野外適存度,野外適存度維持基因重複多樣性!
回顧一下我們研究的兩個品系:品系 MT 含有三個 BCMA 基因,另一個品系 CO 失去了三個 BCMA基因中的 BCMA1(圖二),因此造成了硫代葡萄糖苷的種類差異(據親身嘗試的勇者描述,葉子啃起來真的有差)。
研究人員發現硫代葡萄糖苷在實驗室及野外都能抵抗草食昆蟲啃食植株。利用人為雜交,我們更發現在某些環境,帶有 BCMA1 基因的個體較抗蟲,而在其他環境,卻是不含 BCMA1 的個體較抗蟲,可能是因為每個地點的昆蟲種類不同[2]。因此,環境差異導致草食昆蟲相不同,進而對能夠在當地存活的硫代葡萄糖苷種類進行選擇,最後導致 BCMA1 基因有無及「芥末口味」分布的地理差異(圖二)。
另外,在排除了抗蟲咬的效用後,研究人員發現 BCMA1 的有無還是會影響到野外存活率!這暗示了 BCMA 基因能藉由其他因素影響植物生存:沒有 BCMA1 基因的這些個體(產出較低比例的BC GS),在極端乾旱環境下的存活率會比含有 BCMA1 基因的個體來得好。透過地景生態學大尺度分析,我們更發現採集自較乾旱氣候帶的品系,在溫室中都會產出較低比例的 BC GS,與實驗結果相符。
在這個狀況下,環境的差異會主動去「維持」遺傳及性狀在整個地景尺度的多樣性:因為每個地點都有最適合的硫代葡萄糖苷化學組成,所以沒有任何一個基因型可以獨佔所有環境。這種由不同天擇壓力主動去維持遺傳多樣性的狀況,就被稱為平衡型天擇(balancing selection)。
因此,產出 Met GS 的個體較抗旱,也在某些環境較抗蟲,而產出 BC GS 的個體則是在其他環境較抗蟲。平衡型天擇不只透過對不同類群昆蟲的抵抗力,還會藉由抗旱程度維持 BCMA 基因們在不同野外環境間的多樣性!
瘋狂的基因重複、突變、與丟失造就更多的多樣性
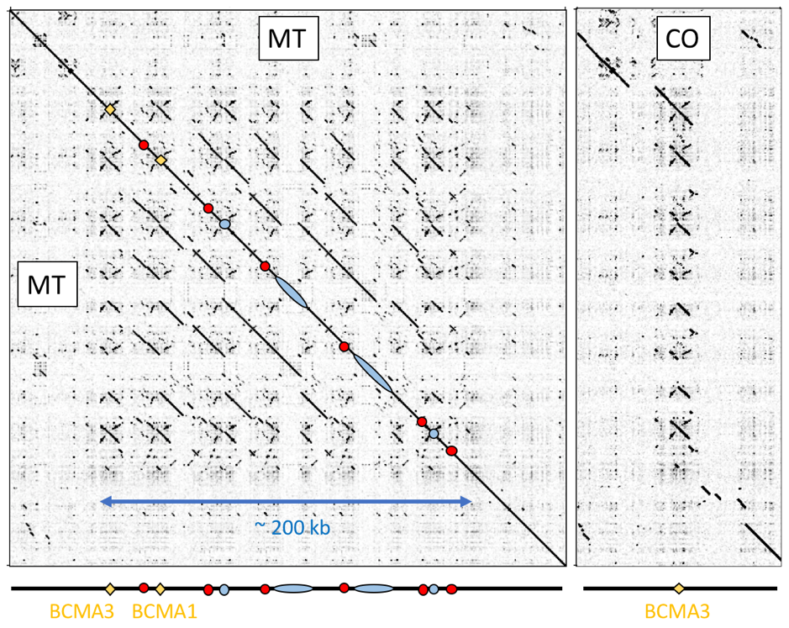
這個故事還沒結束!在七號染色體上的這個位置,MT 品系具有 BCMA1 與 BCMA3 兩個有功能的基因,而 CO 品系缺了 BCMA1(圖二)。但這就是故事的全貌嗎?
透過第三代定序科技,我們重建了兩個品系這個富含重複序列的染色體片段(圖三):MT 品系在這個地方發生了瘋狂的基因重複,把這個位置擴展到超過二十萬個鹼基對,含有起碼 12 個 BCMA 基因!這 12 個之中,其實僅剩原本的 BCMA1 與 BCMA3 具有正常功能,剩下的不是因為片段缺失後只剩一小部分,就是因為中間有轉位子(transposon)的插入讓它無法生成正常蛋白質產物。
換言之雖然有 12 個基因,絕大部分都已經死掉了,只剩兩個有功能。相較之下 CO 品系算是很客氣,在這個區段僅有 BCMA3 一個基因。
這兩個品系的 BCMA 基因數量其實並不特別:透過分析超過 500 個個體的基因體序列,我們推估他們體內的 BCMA 基因數量從兩個到十多個都有。當然這些具有十多個 BCMA 基因的個體,他們體內的 BCMA 有多少還活著,有待進一步研究。有趣的是,我們發現「偏好分佈在潮濕環境的族群」(MT 品系就屬於此族群),體內 BCMA 基因數量比其他族群多很多,會不會其實就跟當地水分含量有關呢?關於天擇與環境如何影響本物種不同族群間的愛恨情仇,請見本團隊近期的另一篇研究。
生物演化不只依靠單一點突變!
演化生物學的一大重點,是探究生物多樣性是怎麼產生的:一個新的表現型怎麼產生?之後有沒有辦法回到原本的祖先型?是經由同一個基因的逆向突變,或是其他因素?
一般概念認為,遺傳改變藉由單一點突變(point mutation)產生。比如某個 DNA 位點的核苷酸 A 會讓植物長出白花,突變為 G 後讓花色變紅。這時如果要反轉性狀,讓花色再度變白,只能期望自然突變再度發生在同一個位點讓 G 變回 A。這樣的機率實在太低了(當然還有其他可能性,先略過不談)。相對地,自然發生的基因重複可以有效地將基因「模組化」,快速地透過大量拷貝貼上、微調、與刪除產生多樣性,不需要慢慢等待點突變再次發生在同一個位點。
相信藉由這個故事,各位可以體認到不斷出現的基因重複、功能改變、及基因缺失也是個驅動演化的重要因子:生物的基因體並不會恆久不變,而是個不斷在突變、天擇、隨機改變、與族群交流之間的動態平衡。
參考文獻
- Prasad et al. 2012. A Gain-of-Function Polymorphism Controlling Complex Traits and Fitness in Nature. Science 337:1081-1083
- Carley et al. 2021. Ecological factors influence balancing selection on a complex trait locus. Nature Ecology & Evolution doi.org/10.1038/s41559-021-01486-0
- Lin et al. 2021. The ecological, genetic, and genomic architecture of local adaptation and population differentiation in Boechera stricta. Proceedings of the Royal Society B 288:20202472
本計劃蒙科技部年輕學者養成計劃之哥倫布計畫補助,特此致謝。






































