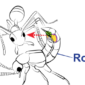
講到新種就好讓人興奮!雖然圖中的菊虎看起來只像是「觸角又長又不發光的螢火蟲」(咦?怎麼聽起來跟蟑螂也有點像?),但卻有好帥氣的原住民名字,牠們是新種菊虎:賽德克狹胸菊虎和鄒狹胸菊虎。
菊虎科(Cantharidae)是一群色彩斑斕、體態修長且翅鞘柔軟的陸生甲蟲。臺灣的菊虎已知有165種並複分3個亞種,其分類學研究始於二十世紀初,主要由歐洲學者發表了多數的種類,至今國內尚未有專家針對此類群進行專門研究。

隱翅菊虎族是一群外型奇特的菊虎類群,牠們的腹部細長且翅鞘短小而無法完全覆蓋後翅。黑足隱翅菊虎是臺灣春夏季十分常見的隱翅菊虎,牠們喜歡聚集於殼斗科花叢間訪花,具有相當好的飛行能力。
由於菊虎的體色變異範圍非常大,所以很難從牠們的外表來鑑定種類。直到20世記中葉,堪稱菊虎分類之祖的Wittmer建立了以雄蟲外生殖器結構作為種類劃分的診斷性特徵後,全球菊虎科分類研究才開始有了系統性的發展。那為何要這麼害羞的以雄蟲的外生殖器結構作為物種定界的依據?
這跟昆蟲學中的鎖鑰假說(lock-and-key hypothesis)有關。這是一種解釋昆蟲種間機械隔離的學說,認為昆蟲的雌、雄兩性的生殖器的相應如鎖與鑰匙般吻合,不同種間的關係明顯不同,走錯家門、帶錯鑰匙可是不得其門而入。
臺灣的地形變化多樣,除了地形隔離外,生物本身也因生態習性而導致的播遷能力不佳,長時間下來即使外表上變異不大,卻產生了物種分化。狹胸菊虎屬(Stenothemus)有別於多數喜於春季現身的菊虎,出沒夏末至冬季的中高海拔森林,夜晚具有趨光性,會被水銀燈的燈光吸引,臺灣本屬成員已知有四種,是筆者非常有興趣的類群之一。

單爪菊虎由於其爪子不開裂或基部不具齒凸而得名,粗腿單爪菊虎是臺灣產單爪菊虎中常見的種類,雄蟲的腿節膨大,雌蟲則否(圖中為雌蟲),白天會訪花而夜晚則具有趨光性。
2013年寒風刺骨的11月天,筆者與學長姐一同前往南投仁愛鄉中高海拔山區採集,一路上各種不順、意外連連,例如才剛出門筆者就把採集用具忘在搭車處的椅子上(等發現的時候人已經在海拔2100公尺的地方了……)以及臨時無法在預訂地點進行燈光誘集的窘境等等諸如此類,最後筆者隨意指向的一處路邊當採集點(就決定是這裡了!!),結果意外地在一群大和田氏狹胸菊虎(Stenothemus owadai)中發現一種外觀上與其他已知種略有差異的狹胸菊虎。
直覺告訴我這可能是個尚未被描述的種類,回到實驗室後,筆者立刻開始進行雄蟲外生殖器的解剖以及比對,當那個擁有特殊幾何結構的外生殖器就這樣「啵!」的拉出來後,已經快要貼在目鏡睡著的我瞬間眼睛亮了,外生殖器結構跟已知種有著顯著的差異,證實這是一種新種的狹胸菊虎。
興奮之餘開始著手進行新種的發表,然而在2014年初夏一個意外的契機,筆者向農業試驗所和倫敦自然史博物館商借到一批產地為南投信義鄉、嘉義阿里山鄉,與筆者採集到的這種非常相似的標本,原來以為是同一種,沒想到檢查雄蟲生殖器結構後,竟然又是一另一個獨立的新種。由於兩個種類的模式產地與賽德克族和鄒族的世居地重疊,便以兩個臺灣原住民族命名為賽德克狹胸菊虎(Stenothemus seediq)、鄒狹胸菊虎(Stenothemus cou)。
- 此研究成果於2015年3月25日,發表於國際期刊【動物分類群】(Zootaxa)。
- 此文由國立臺灣大學昆蟲學系大三生 蕭昀撰寫,響應PanSci 「自己的研究自己分享」,以增進眾人對基礎科學研究的了解。
- 參考文獻:Hsiao, Y (2015) Description of two new species of the genus Stenothemus from Taiwan (Coleoptera: Cantharidae). Zootaxa 3937: 386–392. doi: 10.11646/zootaxa.3937.2.9