會寫這篇文章的原因是一覺醒來,發現臉書上的朋友突然對「中度干擾假說」充滿好奇。不禁嘖嘖稱奇,這個生態學的假說,還有機會在鄉民之間熱絡起來?不過無妨,就藉由這個機會介紹一下這個假說是在幹嘛,沒事多喝水、沒事長知識。不過先說明一下,這個玩意兒要用一些其他的專有名詞來解釋,如果每個名詞都講清楚篇幅會過長,所以請大家自行估狗或有空再聊聊其他的專有名詞吧。
研究生物多樣性的時候,常常是在問一些問題:「哪裡生物的種類多?哪裡生物的種類少?為何而多?為何而少?」。當然,影響一個地區物種多寡的原因很多,其中一個主題是探討多樣性(diversity)和干擾(disturbance)之間的互動關係。而中度干擾假說(intermediate disturbance hypothesis, IDH)就是其中一個解釋。
有事沒事戳一下會怎樣?
在生態學中,干擾(disturbance)是一個暫時使環境狀態發生改變的事件,這個改變幅度超過環境平時的小幅變動。干擾的影響通常劇烈且快速,會導致部分生物消失。不同的干擾作用的空間範圍與時間長短也不盡相同,規模可大可小,頻率孰繁孰罕,天然的干擾如火災、水患、颱風、地震、海嘯、氣候變遷和昆蟲大發生,人類的干擾則包括過度開發及過度獵捕等等[1]。這樣看起來,干擾似乎不是什麼好東西,無論規模大小,感覺影響挺負面的。換句話說,干擾的強度或頻度超過某個程度,會削弱生物的繁殖與擴張,使當地的群集不飽和,物種多樣度沒有達到應有的水準。
戳幾下似乎比較好?
然而,也有觀點認為干擾是維繫生物多樣性的重要元素[2,3]。在探討多樣性和干擾之間的關係時,研究珊瑚和樹木等固著性生物(sessile species)的案例中[2]發現:使物種多樣度(species diversity)最高並非發生在干擾的強度(或頻度為免版面混亂,看到強度請自行加上「或頻度」)最高或最低時,而是在不高不低的中度(並非指中數或平均值)。主張中度干擾假說的論述認為:在干擾強度太強時,導致大多數的物種難以生存,僅有少數繁殖力強或擴張力強的拓殖者(colonizer)能夠生存,如老鼠和禾草。干擾強度太低時,優勢的競爭者(competitor)會極度強盛,競爭排斥較劣勢的物種,物種多樣度也不高。因此,只有在適度的干擾之下,拓殖者和競爭者能夠共存,因而有最高的物種多樣度,使干擾和多樣性之間關係呈現駝峰狀關係(hump-shaped relationship)。
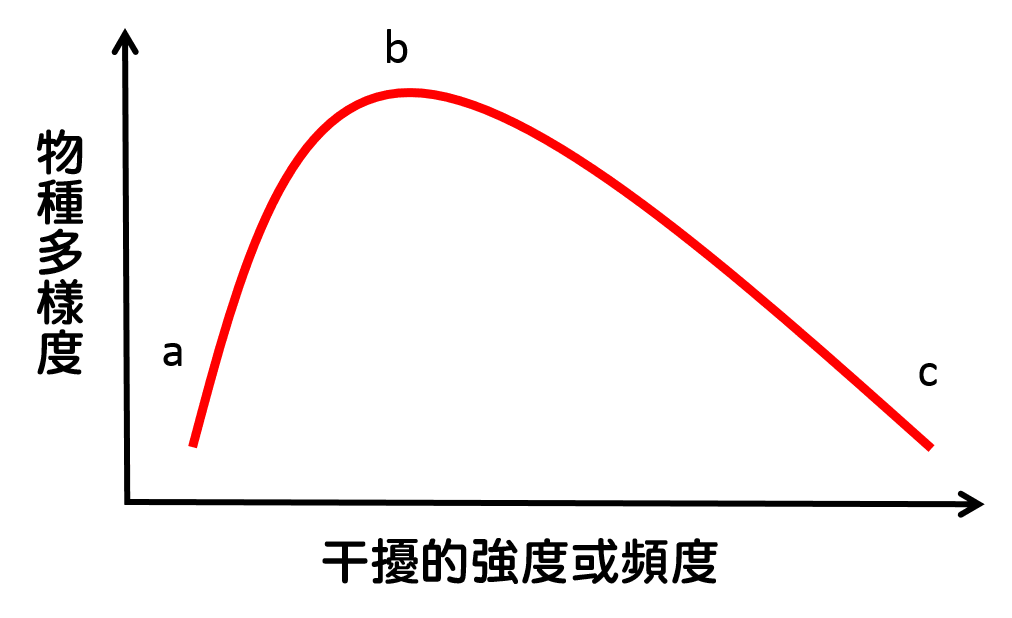
中度干擾假說的知名研究案例是研究潮間帶礫石上的附生藻類[4]。經常受海浪拍打而滾動的礫石體積較小,反之則體積較大,作者以此作為受干擾程度的指標。作者發現:(1)在較小顆、較常滾動的礫石上,藻類的物種多樣度較低,主要由生長快速的拓殖者組成。(2)大顆的礫石上則主要由優勢的競爭者所組成,多樣度也較低。(3)不大不小的礫石上同時具有競爭者和拓殖者,呈現較高的物種多樣度。
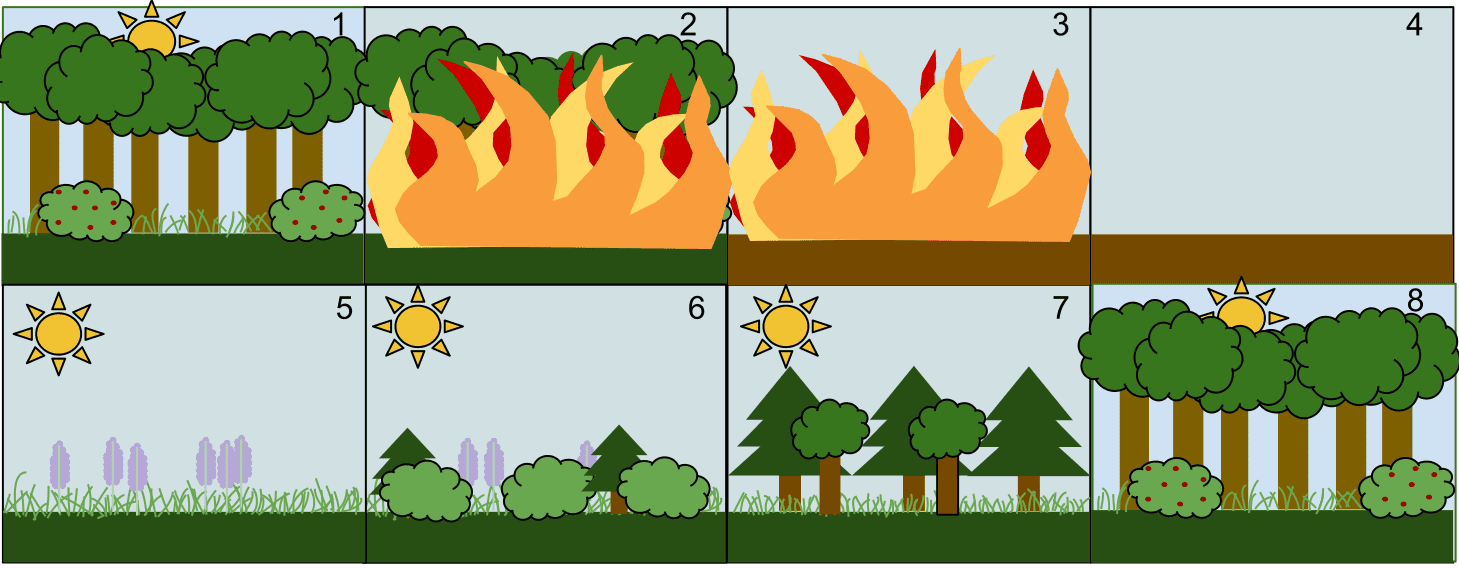
另一方面,是從地景生態學(landscape ecology)的觀點來看中度干擾假說:干擾會使部分棲地及其中生物消失,使當地的生物進行次級演替(secondary succession),可以說是刺激當地生物重新拓殖的開始。無論規模大小的干擾發生,都會改變當地的地景,形成各種不同面積的小區塊,這種區塊稱為「孔隙」(gap)。例如森林中間發生了小火災,燒了一小部分,這個部分就是孔隙。接著,孔隙中的小草、灌木、小樹、大樹會隨著不同的時期慢慢長回來(如下圖5, 6, 7, 8),這些不同的階段就稱為演替階段(succession stage)。
所以,以植物為例的話,各位可以看到下圖,不同演替階段的植物成員都不盡相同。也就是說,中度的干擾,前前後後在森林裡形成許多孔隙,因為孔隙形成的時間不同,隨著時間過去,每一個孔隙所處的演替階段也不同。換句話說,這個森林裡面含有的各種不同演替階段,使整體的空間異質度(spatial heterogeneity, 指空間複雜的程度)較高,所含的微棲地(micro-habitat)也就更多樣,能容納不同生態棲位(ecological niche)的生物也越多,使物種多樣度增加。這樣的理論,稱為「空間異質度理論」(spatial heterogeneity theory)[5]。生態棲位越多,越能避免因種間競爭(interspecies competition)所造成的局部絕種。
中度干擾假說受到的批評
那麼,干擾和物種多樣度的關係都像中度干擾假說的預測一樣,呈現駝峰狀的關係嗎?肯定不是這樣的啦,回顧87篇研究干擾與物種多樣度關係的文獻,發現有25篇認為兩者之間無關係、25篇呈現正向關係、16篇為駝峰狀關係、18篇為負向關係、3篇為倒U型關係[6]。更近期的回顧也顯示駝峰狀關係並不普遍[7]。競爭排斥原理(competitive exclusion principle)不是只會導致劣勢的物種滅絕,也有可能促成物種的種化(speciation)。對具備移動能力的動物而言,大可一走了之,特別是在棲地異質度高的環境,另覓生存新天地不見得是難事[8, 9]。有的觀點認為,干擾並非作用於降低競爭排斥的力度,而是使生物的豐度(abundance)下降之後,造成平均個體的成長率(per capita growth rate)呈現負值,使族群一蹶不振[10]。另外,有些觀點則認為干擾和物種多樣度之間會比測互相影響、互為因果,兩者無限循環的交互作用。當我們觀察現象時,已經作用許久,難以釐清究竟是雞生蛋還是蛋生雞[11]。

之所以會有這些爭議,通常是兩者所見的環境狀況、生物種類、空間與時間的尺度有所不同而導致的歧見。干擾對生物的影響,要考量干擾作用的空間尺度與觀察空間尺度的差別。如果干擾的空間尺度大於所觀察的空間尺度,例如一顆巨石砸毀整個花盆,較適合以干擾作用解釋。如果干擾的空間尺度遠小於所觀察的空間尺度,例如一大片森林遇到多次小規模的崩塌和火災,可能較適用中度干擾假說的解釋。
大自然的干擾與生物群集之間,經過頻繁且複雜交互作用,形成我們現今所見的生命世界,而且持續的在變化。雖然中度干擾假說的適用性有限,甚至這個理論該不該存在,學術界正吵得火熱[10, 12, 13]。但是,至少我們很確定,過強的干擾會讓生命大量的消失。以往是自然發生的干擾與生物互動,現在還增加了許多種類與強度不盡相同的人為干擾,對生命世界帶來龐大的衝擊。如果我們還無法控制這些過度的人為干擾,一但超出生物所能負荷的臨界,那麼這個生命世界也只能向下沉淪。
引用文獻
- 1. Grime, J. P. 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist, 111: 1169-1194.
- 2. Connell, J. H. 1978. Diversity in tropical rain forests and coral reefs. Science, 199: 1302-1310.
- 3. Huston, M. 1979. A general hypothesis of species diversity. American Naturalist, 113: 81-101.
- 4. Sousa, W. P. 1979. Distubance in marine intertidal boulder fields: the nonequilibrium maintenance of species diversity. Ecology, 60: 1225-1239.
- 5. MacArthur, R. H. and J. W. MacArthur. 1961. On bird species diversity. Ecology, 42: 594-598.
- 6. Mackey, R. L. and D. J.Currie. 2001. The diversity-disturbance relationship: is it generally strong and peaked? Ecology, 82: 3479-3492.
- 7. Hughes, A. R. et al. Reciprocal relationships and potential feedbacks between biodiversity and disturbance. Ecology Letters, 10: 849-864.
- 8. Bruno, J. F. et al. 2003. Inclusion of facilitation into ecological theory. Trends in Ecology and Evolution, 18: 119-125.
- 9. Kimbro, D. L. and E. D. Grosholz. 2006. Disturbance influences oyster community richness and evenness, but not diversity. Ecology, 87: 2378-2388.
- 10. Fox, J. W. 2013. The intermediate disturbance hypothesis should be abandoned. Trends in Ecology and Evolution, 28: 86-92.
- 11. Hughes, A. 2010. Disturbance and Diversity: An Ecological Chicken and Egg Problem. Nature Education Knowledge, 3(10): 48.
- 12. Sheil, D. and D. F. R. P. Burslem. 2013. Defining and defending Connell’s intermediate disturbance hypothesis: a response to Fox. Trends in Ecology and Evolution, 28(10): 571-572.
- 13. Fox, J. W. 2013. The intermediate disturbance hypothesis is broadly defined, substantive issues are key: A reply to Sheil and Burslem. Trends in Ecology and Evolution, 28(10): 572-573.
延伸閱讀
- 超級反對中度干擾假說的Jeremy Fox的部落格,他認為這假說根本是個打不死的殭屍,恨不得把它給廢了。這個部落格文章戰意甚深,底下的回應也很精采,有興趣的讀者不妨看看,自己跳下去戰也行。