- 本文轉載自特有生物研究保育中心,《自然保育季刊》第 112 期
- 作者 / 陳子浩|行政院農業委員會特有生物研究保育中心計畫助理)、林彥博|行政院農業委員會特有生物研究保育中心助理研究員
特生中心團隊長達 8 年的生態監測調查,讓我們知曉蘇花改公路令人驚嘆的生物多樣性。上一篇〈蘇花公路的兩爬生物點點名!(上)——蘇花改特輯(三)〉介紹了可愛的兩棲蛙類和蜥蜴大家庭,接下來繼續瞭解擁有多樣生活型態與食性的蛇類,以及劃分生物地理區界線的清水斷崖!
生活形態與食性各異的蛇類
蛇類的神出鬼沒眾所皆知,這也增加了調查的難度。森林邊緣、開闊或墾地、森林底層和水域周遭都可能與牠們不期而遇,除了透過定點穿越線調查,路殺調查有助瞭解蘇花改沿線的蛇類相。

目前調查到的蛇類裡茶斑蛇、紅竹蛇、赤背松柏根主要在森林底層活動。為臺灣四大美蛇之一的紅竹蛇喜好捕食小型哺乳類,而赤背松柏根為卵食性,茶斑蛇則仰賴小型兩棲爬行動物為生,食性明顯不同。

在森林邊緣活動的物種有善於夜襲休息鳥類或爬行類的大頭蛇,專食陸生軟體動物的泰雅鈍頭蛇,伏擊小型哺乳類的龜殼花,日間靠著火眼金睛四處游獵兩棲爬行和魚類的過山刀, 還有臺灣三種綠色蛇類之一的青蛇。大型的細紋南蛇、南蛇、王錦蛇和眼鏡蛇較能適應人為干擾環境,常於墾地或鄰近住家的開闊地活動,捕食小型哺乳類、兩棲爬行類為食,其中眼鏡蛇因地理隔離產生遺傳分化,使得東部地區族群在外部形態上有著黑色的腹鱗特徵 (Lin et al. 2008)。

此外,水域周遭環境也能發現不少蛇類棲息,在進行夜間兩棲類調查時,常會看見赤尾青竹絲攀附在水邊的灌叢上,坐等獵物進入攻擊範圍;雨傘節和紅斑蛇會在水邊的草叢游獵,草花蛇則在水田或草澤捕魚獵蛙,是一種高度仰賴靜水域的蛇種,在臺灣西半部較為常見,但近年來水田比例下降與農藥使用而造成棲地劣化,使其族群大不如前,如今已然成了臺灣保育類蛇種。

清水斷崖劃分生物地理的區界線
臺灣島位於亞洲大陸東南方,東邊與南邊分別與琉球島弧及呂宋島弧相連接。如此特殊的地理位置使得臺灣的生物相具有複雜的組成來源,又加上臺灣島在歐亞大陸及菲律賓板塊的擠壓下地形異質性相當高,有高山、丘陵、淺山及平原地區,可提供各式各樣的生物生活其中,因此臺灣是在研究生物地理學一個很適合的熱點區域。
臺灣島上生物的分化模式大致可分成三個模式:
- 在冰河時期,亞洲大陸的祖先族群藉著海平面下降形成陸橋,一次或多次到達臺灣,而後冰河期結束後形成地理隔離而分化。
- 由鄰近島嶼上的生物經過天然途徑傳播到臺灣島上落地生根,形成在遺傳組成上具有獨特性的族群。
- 因為臺灣的高異質性的地理結構造成不同族群的生殖隔離而在島內分化成不同物種 (陳玉峰 1995;鄧書麟及呂福原 2015; Tseng et al. 2015)。

臺灣的兩棲爬行類動物中也有種化事件發生,例如雪山草蜥、翠斑草蜥及鹿野草蜥的島內種化事件 (Tseng et al. 2015)。其中翠斑草蜥及鹿野草蜥僅是因為狹窄的立霧溪阻隔而產生物種分化事件。而從立霧溪口延伸至清水斷崖,即大約是在崇德至和仁之間,這段 5-6 km 長的狹長地帶,一邊是山,一邊是海,斷崖地形幾乎垂直插入太平洋的山壁,可能是臺灣最壯麗的地理景觀之一。但對於臺灣的兩棲爬行類而言, 清水斷崖卻也是個難以跨越的地理屏障,使得該處在臺灣北部及東部的生物地理區界線上扮演一個重要的角色。 除了翠斑草蜥與鹿野草蜥外,有許多兩棲爬行類的演化系群分化或是分布界線都座落在清水斷崖附近,例如褐樹蛙在清水斷崖兩側的族群遺傳組成上有非常大的差異 (Lin et al.2012)。在臺灣兩棲爬行類研究中常有提到在其分布界線上,諸如面天樹蛙、黃口攀蜥、福建大頭蛙等物種只分布於清水斷崖以北的區域,而半葉趾虎、鱗趾虎及梭德氏草蜥只分布於清水斷崖以南的區域。

在我們過去8年多在蘇花公路沿線所進行的兩棲爬行類多樣性調查研究中,黃口攀蜥分布南限為東澳地區,福建大頭蛙是蘇澳地區,以上2種兩爬物種的發現紀錄的確在過了清水斷崖以南區域即消失了,面天樹蛙則分布到崇德(清水斷崖以南)。以往文獻紀載中分布於西南及東南部、蘭嶼及綠島的半葉趾虎和鱗趾虎,這幾年間陸續於清水斷崖以北的區域被記錄,半葉趾虎從和仁、和中,最北分布至南澳,而和仁、和中地區也有發現鱗趾虎的紀錄,諸多研究結果顯示清水斷崖除了讓人們南北往來交通受到不小的阻礙之外,對於兩棲爬行類的物種分化及多樣性也具有令人驚奇的影響。

兩爬類的悲歌——蘇花公路沿線的路殺現象
道路的開發與建設往往會衝擊到野生動物的原始生存環境,因對環境變動較為敏感及其特殊的生態特性,兩棲爬行類受到道路開發的影響甚劇,尤其是道路致死 (也稱為路殺) 問題。我們在蘇花公路沿線記錄到的路殺兩棲爬行動物紀錄中,爬行類有 115 筆, 路殺隻數前三高分別為青蛇 41 筆、 雨傘節 18 筆及龜殼花 11 筆; 兩棲類有 53 筆,隻數前三高分別為盤古蟾蜍 13 筆、 拉都希氏赤蛙 11 筆及黑眶蟾蜍10 筆。單就數據上來看爬行類的路殺問題似乎比較嚴重,但其實不然,較小型的兩棲類的肌肉量較少,容易因車輛重複輾壓而支離破碎,快速地腐化殆盡,或因其他動物的撿食而消失,因此較不容易被發現記錄。
兩棲類經常於陸域與水域之間移動,能使用的水域類型相當廣泛,包含路邊的臨時性水灘或溝渠,而路燈下聚集的昆蟲有時也會吸引牠們聚集捕食,大幅提高兩棲類在馬路上活動的機會,如盤古蟾蜍和黑眶蟾蜍的路死個體即常被發現於路燈附近的道路上。蛇類則因喜愛夜間馬路上殘留的熱能,加上尋覓獵物時常穿越馬路,身體修長閃躲不易而常慘死輪下。其中青蛇在南澳地區的路殺數量記錄相當多,推測可能為南澳地區森林相較完整,是青蛇良好的棲地;此外青蛇為日行性蛇類,日落後會再回到樹上休息, 活動時間與人類重疊度高,這種在覓食及休息場域間移動的行為,往往使其在穿越道路時遭輾死亡。

道路致死的調查,除了有助於彌補兩爬動物相調查不易的困難,尚能藉由資料點位統整找出路殺熱點,並探討路殺主因,配合施工單位改善、新增道路設施,如設計不良的水溝、邊坡、護欄或增加生物廊道等,有機會降低棲地切割帶來的路殺壓力,在保護動物的宗旨下同時提升行車安全而達到雙贏的效果。
保育珍貴生物多樣性刻不容緩
兩棲爬行動物因為豐富的生活史多樣性及位居重要的生態鏈地位,相當適合作為一個自然環境的指標物種。但這兩大類群當前正面臨著巨大的威脅,除了棲地退化與消失造成的族群數量下降外,棲地破碎化與路殺的問題也隨著人為活動而日漸嚴重,長期以來人們對於兩棲爬行類認知不足而造成的誤解也使得兩棲爬行類的保育之路走的更為艱辛,除了強化民眾基礎知識與觀念的建立,利用廣大公民科學家的力量,蒐集兩棲爬行類的基礎生態資料或建置物種分布資料庫,將可用於生態永續工程開發或減少道路建設對野生動物的影響。
蘇花改公路已於 2020 年 1 月 6 日全線通車,研究人員持續監測蘇花改沿線兩棲爬行動物族群以瞭解其回復的狀況,除了盼望物種能盡速回復至原來的棲地外,同時亦嘗試找出蘇花改公路的路殺熱點,並向施工單位提出建議用對野生動物友善的工程方式,將有助於降低路殺帶給生物族群的生態衝擊,以維護當地的生物多樣性。
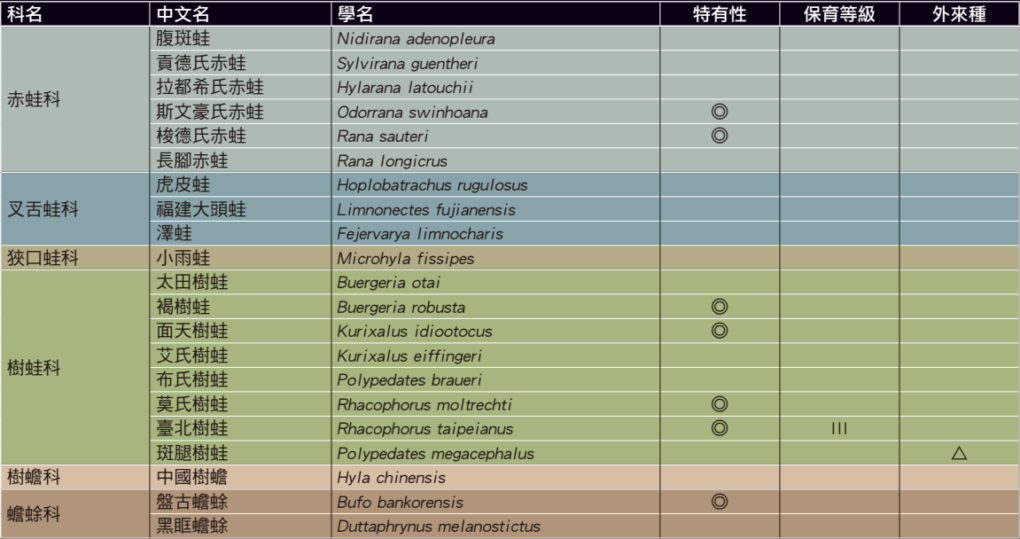
注 – ◎:臺灣特有種 ○:臺灣特有亞種 △:外來入侵種 I:瀕臨絕種野生動物 III:其他應予保育之野生動物。圖/《自然保育季刊》第 112 期
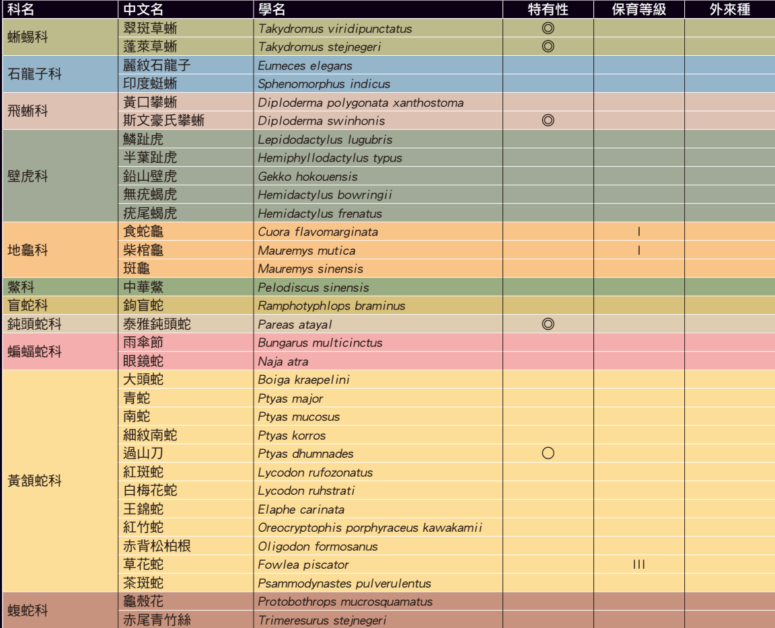
注 – ◎:臺灣特有種 ○:臺灣特有亞種 △:外來入侵種 I:瀕臨絕種野生動物 III:其他應予保育之野生動物。圖/《自然保育季刊》第 112 期