


- 本文轉載自特有生物研究保育中心,《自然保育季刊》第 112 期
- 作者 / 鄭錫奇|行政院農業委員會特有生物研究保育中心研究員兼主任秘書、陳宏彰|行政院農業委員會特有生物研究保育中心計畫助理、周政翰|台灣蝙蝠學會理事
一條安全回家的路
政府呼應民意、承諾積極改善蘇花公路行車路況,給國人一條安全回家的路,交通部公路總局蘇花公路改善工程處(下簡稱蘇改處)歷時 9 年的努力,全長約 38.8km 的「台 9 線蘇花公路山區路段改善工程」(下簡稱蘇花改)終於在 2020 年 1 月 6 日全線通車。此工程包括 8 座隧道、許多橋梁與部分路堤, 總經費新臺幣 551 億元。

所謂「蘇花改」是指改善原蘇花公路蘇澳到東澳、南澳到和平、和中到大清水部分山區的險峻路段,完工後蘇花公路大幅縮短 18.65km,在兼具行車安全與舒適之餘,往返宜蘭與花蓮之間於正常時刻可省下逾 1 小時的行車時間。
蘇改處在執行改善工程之初,即依據蘇花改主要環評決議事項,於 2010 年 11 月 10 日以行政協助委託行政院農業委員會特有生物研究保育中心(下簡稱特生中心)執行「台 9 線蘇花公路山區路段改善計畫(蘇澳~東澳、南澳~和平、和中~大清水)施工中暨營運階段指標生物研究計畫」,總計畫包含了 10 項子計畫,其中一項為「翼手目動物研究調查計畫」。
近年政府進行重大建設時日益重視工程對環境及生物多樣性的影響評估,蝙蝠類因其物種繁多、族群數量龐大、食性獨特、活動範圍廣泛且對環境變化敏感,其生存棲地品質攸關族群存續狀況,因此相當適合作為反應環境變化的類群。雖然行政院環保署於 2011 年 7 月修訂公告的「動物生態評估技術規範」始增列蝙蝠類群於陸域哺乳動物調查與監測中,但蘇改處仍超前部署,規劃在台 9 線蘇花改道路施工範圍及周邊區域進行全線的翼手目(蝙蝠類)調查,藉以評估該類群是否會受到蘇花改工程的影響,並探討如何藉由所得資料降低工程衝擊,以保有在未來完工後復原的潛力。因此,讓我們有幸一探蘇花公路沿線的蝙蝠多樣性。

神奇的飛行哺乳類
世界上已知的蝙蝠種類超過 1,400 種 (Simmons 2019),除了南、北極地外,幾乎廣泛分布在每個地區。蝙蝠是唯一具飛行能力的哺乳動物,生存在地球上至少已有 5,200 萬年 (Simmons et al. 2008),迄今演化出高度多樣性的物種與許多特殊的生存本領,除了飛行能力(雙翼)外,尚有變溫調節生理(休眠與冬眠)機制、多數以超音波偵測 (概分定頻 CF 與調頻 FM 型式)搜尋與覓食,以及生殖生理調適(授精、著床或發育延遲)等。
研究得知,蝙蝠在生存的生態系中扮演維繫平衡與穩定發展的角色,尤其藉由食性所提供的重要生態功能,包括抑制昆蟲 (insect suppression,如農業害蟲)、傳播種子 (seed dispersal)、傳花授粉 (pollination) 等 (Justin et al . 2014)。族群數量龐大的蝙蝠對人類農作物的經濟收成有顯著的影響,Bea 等人 (2013) 研究發現當食蟲性蝙蝠和鳥類存在於農作區取食昆蟲,人類所種植的作物可可亞樹 (cacao trees) 收成可增加達 31%。
臺灣蝙蝠知多少?
臺灣蝙蝠最早的紀錄為知名博物學者史溫侯 (Robert Swinhoe) 在 1862 年對游離尾蝠及一種鼠耳蝠的描述。之後歷經歐、美及日本等諸多學者的努力,至 1950 年代已 大致建立了 19 種臺灣蝙蝠的名錄資料 (陳兼善 1956)。

近代另一波臺灣蝙蝠物種的採集與分類研究則由日本學者吉行端子 (M. Yoshiyuki) 在 1991 年發表了臺灣長耳蝠 (Plecotus taivanus) 開啟,隨著調查器材與研究技術的精進,許多臺灣的新種、新紀錄種或分類地位再確認種陸續被釐清發表,譬如長相特殊的管鼻蝠屬 (Murina spp.) 類群 (Kuo et al. 2006、2009)、離島馬祖的臺灣新紀錄種灰伏翼 (Hypsugo pulveratus)(張簡琳玟等 2013)、物種繁多的鼠耳蝠類群 (Myotis spp.)(Csorba et al. 2014; Ruedi et al. 2015)、喜隱棲在芭蕉捲葉的玄彩蝠 (Kuo et al. 2017),以及分布於金門地區的新紀錄種大足鼠耳蝠 (Myotis pilosus)(周政翰等 2019),使得臺灣地區(含金門、馬祖離島)的蝙蝠物種達 37 種。根據「野生動物保育法」之陸域保育類野生動物名錄,臺灣的蝙蝠有兩種列名保育類,即瀕臨絕種的臺灣狐蝠 (Pteropus dasymallus formosus) 和珍貴稀有的臺灣無尾葉鼻蝠 (Coelops frithii formosanus)(林務局 2019)。

而由特生中心出版的《2017 臺灣陸域哺乳類紅皮書名錄》,則將臺灣狐蝠列屬國家極度瀕危 (nationally critically endangered, NCR) 等級、霜毛蝠 (Vespertilio sinensis) 列為國家瀕危 (nationally endangered, NEN) 等級,臺灣無尾葉鼻蝠及金黃鼠耳蝠 (Myotis formosus flavus) 則列為國家易危 (nationally vulnerable, NVU) 等級,為需要特別關注與保育的物種 (鄭錫奇等 2017)。

蘇花公路蝙蝠資料有限
當我們回顧蘇花公路沿線有關蝙蝠的文獻時, 發現當地蝙蝠相關的調查資料極其有限。相關報告多為環評報告,諸如「國道東部公路蘇澳花蓮段工程規劃環境影響說明書」(1998)、「國道東部公路蘇澳花蓮段環境影響差異分析暨環境現況差異分析及對策檢討報告」(2006) 等報告書,均僅記錄東亞家蝠 1 種;弘益生態有限公司於 2010 年執行撰寫的 「台 9 線蘇花公路山區路段改善計畫生態環境調查報告書」中則列有 3 科 7 屬 7 種蝙蝠,包括臺灣葉鼻蝠、臺灣大蹄鼻蝠、臺灣小蹄鼻蝠、東亞家蝠、高頭蝠、黃頸蝠及摺翅蝠(現稱東亞摺翅蝠)。
2011 年則有「理新礦業之台濟採字第 5061 號申請核定及變更核定礦業用地環境影響說明書」,曾於花蓮縣壽豐鄉荖腦山北方之山谷(屬木瓜山事業區第 97 林班地)進行探勘,並無發現任何蝙蝠;福邦工程顧問有限公司 (2011) 進行嘉新水泥股份有限公司台濟採字 第 5454 號礦區申請核定及變更核定礦業用地計畫和平礦場開發計畫時,針對花蓮縣秀林鄉和平村,以及宜蘭縣南澳鄉與花蓮縣秀林鄉交界之和平溪 (大濁 水溪) 中、下游北岸山麓開發案進行生態調查時,亦僅列出東亞家蝠 1 種蝙蝠。
若擴大地理尺度,特生中心研究人員曾在 2003-2004 年間於宜蘭縣和花蓮縣進行哺乳類動物資源調查,累積發現了至少 4 科 12 屬 20 種蝙蝠 (鄭錫奇與張簡琳玟 2003,鄭錫奇等 2004),然而該調查報告中樣點所涵蓋的海拔範圍及棲地型態相當廣泛,而台 9 線蘇花公路山區路段大致侷限在海拔 500m 以下的場域,因此實際有多少蝙蝠物種及族群在此區段棲息、活動或棲住在哪裡?以及與當地棲地型態的相關性?均所知有限。

運用多樣的調查方法
研究人員在台 9 線蘇花公路沿線及山區路段改善施工範圍選定適當的調查樣區(暨樣線或樣點), 並視工程施工進度、天候因素、地形地貌變動等實際狀況調整。為完整收集蝙蝠類資料,我們以網具捕捉、超音波回聲測錄辨種及棲所勘查等 3 種方法進行調查(參考鄭錫奇與周政翰 2009)。
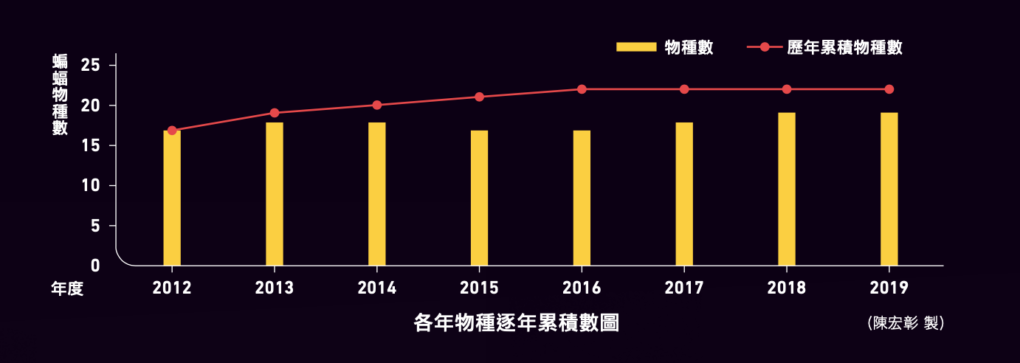
由 2012 至 2019 年度總共在蘇花公路沿線及周邊勘查了 58 個樣區,並選擇其中 53 處設置調查樣區,包括 31 處適合架設網具 (豎琴網為主) 以捕捉夜間活動的蝙蝠、 23 處適合進行超音波的測錄調查,以及 15 個蝙蝠潛在棲所之探查樣區;部分樣區在架網捕捉蝙蝠的 同時亦進行超音波測錄。每個年度按冬季 (12-2 月)、春季 (3-5 月)、夏季 (6-8 月)、秋季 (9-11 月) 來區分為 4 個季節,每季進行 2 次調查。除 2012 年冬季因計畫期程未進行調查外,總計完成了 8 個年度共 31 季的調查工作。
豐碩的調查成果
近 8 個(2012-2019)年度的調查時程,我們總共發現 5 科 14 屬 22 種蝙蝠,包括蹄鼻蝠科 2 種(臺灣大蹄鼻蝠和臺灣小蹄鼻蝠)、葉鼻蝠科 1 種(臺灣葉鼻蝠)、蝙蝠科 17 種(黃頸蝠、堀川氏棕蝠、絨山蝠、東方寬耳蝠、東亞家蝠、山家蝠、臺灣家蝠、寬吻鼠耳蝠、赤黑鼠耳蝠、長趾鼠耳蝠、長尾鼠耳蝠、毛翼管鼻蝠、金芒管鼻蝠、黃胸管鼻蝠、臺灣管鼻蝠、隱姬管鼻蝠及玄彩蝠)、摺翅蝠科 1 種(東亞摺翅蝠),以及游離尾蝠科的東亞游離尾蝠,種類約占 臺灣本島食蟲性蝙蝠(以 32 種計)的 68.75%;其中屬於臺灣特有種者有臺灣大蹄鼻蝠、臺灣小蹄鼻蝠、 黃頸蝠、寬吻鼠耳蝠、長趾鼠耳蝠、黃胸管鼻蝠、 臺灣管鼻蝠及隱姬管鼻蝠等 8 種,臺灣特有亞種者 有臺灣葉鼻蝠及堀川氏棕蝠 2 種(表 1)。

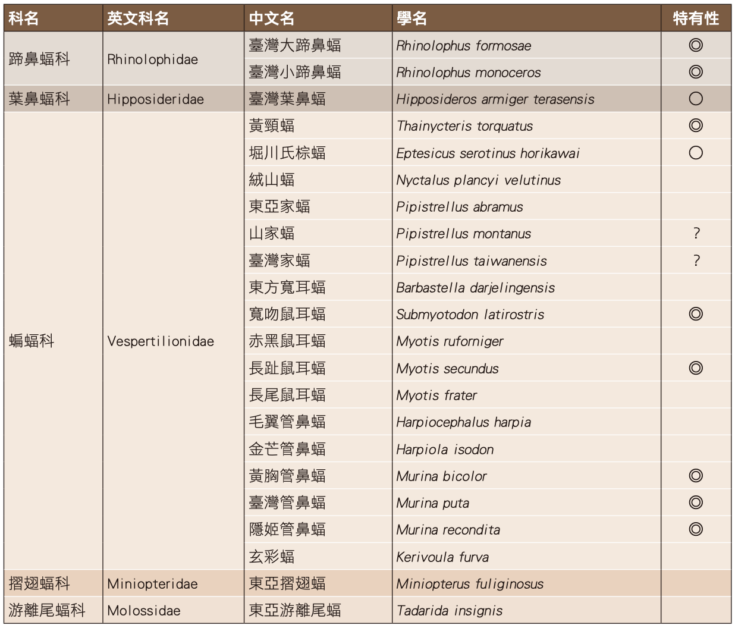
註:至 2019 年止共計 5 科 14 屬 22 種。學名及特有性參考鄭錫奇等 (2017) 及 Wilson and Mittermeier (2019), 其中山家蝠及臺灣家蝠之學名按吳建廷 (2007)。 ◎:臺灣特有種 ○:臺灣特有亞種 ?:特有性未確認。圖/《自然保育季刊》第 112 期
不同調查方法的結果有異
不同的調查方法所獲得的調查結果不盡相同。歷年由網具共捕獲 16 種 568 隻次的蝙蝠,分屬 4 科 11 屬,其中數量最多者為臺灣管鼻蝠(271 隻次),其次為隱姬管鼻蝠(80 隻次)和長趾鼠耳蝠(68 隻次),而臺灣小蹄鼻蝠、臺灣葉鼻蝠、赤黑鼠耳 蝠、長尾鼠耳蝠及玄彩蝠則有 25-38 隻次之捕捉紀錄,至於東亞摺翅蝠、黃胸管鼻蝠、寬吻鼠耳蝠、 金芒管鼻蝠、東方寬耳蝠、山家蝠、臺灣家蝠、毛翼管鼻蝠等則都在 10 隻次以下,相對較少。

由超音波測錄辨種法獲得至少有 17 種蝙蝠在當地活動的資料,其中有效音頻數較多者依序為長趾鼠耳蝠(9,168 筆)、山家蝠(8,481 筆)、堀川氏棕蝠(4,874 筆)及東亞家蝠(1,344 筆),而赤黑鼠耳蝠、 長尾鼠耳蝠、臺灣葉鼻蝠、臺灣小蹄鼻蝠、臺灣大蹄鼻蝠、東亞游離尾蝠、絨山蝠、東亞摺翅蝠及臺灣管鼻蝠等亦有數百筆之多,至於黃頸蝠、臺灣家蝠、玄彩蝠及隱姬管鼻蝠等物種則在 20 筆以下,相對少了很多。

部分物種似乎廣泛分布在台 9 線蘇花公路沿線與周邊區域,但我們僅發現 7 種蝙蝠的棲所,包括喜歡住在洞穴、隧道、涵洞等處的臺灣大蹄鼻蝠、臺灣小蹄鼻蝠、臺灣葉鼻蝠、長趾鼠耳蝠及東亞摺翅蝠,夜間會暫棲在橋墩下的堀川氏棕蝠、臺灣小蹄鼻蝠和臺灣葉鼻蝠,以及偏好棲息在新鮮芭蕉捲葉中的玄彩蝠;其中為確定種類而進行捕撈及標放者有 136 隻次蝙蝠,包括玄彩蝠 88 隻次、堀川氏棕蝠 37 隻次、臺灣大蹄鼻蝠 9 隻次,以及各僅 1 隻次的臺灣葉鼻蝠和長趾鼠耳蝠。

完整資料需多種調查方法

綜合上述 3 種調查方法而獲得的 22 種蝙蝠中, 多數種類可同時由網具捕捉及超音波發現,然而黃頸蝠、堀川氏棕蝠、絨山蝠、東亞家蝠及東亞游離尾蝠等 5 種僅藉由超音波測錄辨種得知,並無網具捕捉紀錄,其中僅堀川氏棕蝠曾被發現其夜間棲息處所(橋墩下)而捕撈確認;於東方寬耳蝠、 寬吻鼠耳蝠、毛翼管鼻蝠、金芒管鼻蝠及黃胸管鼻蝠等 5 種則無測得超音波資料,乃由網具直接捕獲發現。


此外,比較網具捕捉及超音波測錄辨種所得到的優勢物種亦有顯著不同:捕捉調查以臺灣管鼻蝠、隱姬管鼻蝠及長趾鼠耳蝠為數量較多的優勢種,而在超音波測錄則顯示出長趾鼠耳蝠、山家蝠、堀川氏棕蝠及東亞家蝠為活躍種類。其中長趾鼠耳蝠在兩種調查方法皆能有效記錄,然而管鼻蝠類(如臺灣管鼻蝠與隱姬管鼻蝠等)的超音波相對微弱,需在短距離內始能測得,而玄彩蝠音頻甚高(最高可達 280kHz,為臺灣蝙蝠超音波最高者),其往往不易被一般的超音波偵測器(如 Anabat 系統)所錄到。

因此,若單純只採用超音波回聲測錄辨種法調查,可能就無法確切掌握部分物種 的存在;相對的,許多偏好高空或空曠區域飛行覓食的蝙蝠種類(如堀川氏棕蝠、絨山蝠、東亞游離尾蝠或家蝠類群)則不易靠網具捕捉發現,就必須依賴超音波測錄調查輔助證實其存在的證據。
接著閱讀:台灣蝙蝠知多少?沿著蘇花公路,探尋豐富的蝙蝠多樣性(下)——蘇花改特輯(二)
































-200x200.jpg)








{kind=link}