文 / 游崇瑋
好啦,不只是最近當紅的泰雅鈍頭蛇,所有的蛇類都不該被當成食物的理想對象。蛇類並沒有什麼藥用的功效,身為物資相對不缺乏的台灣人,我們有更多的豢養動物可以選擇,畢竟不是每個人都要當貝爾大叔,不是嗎?
一個又一個全神貫注、瘋狂找蛇的夜晚;一次又一次舉著相機、專注細節的快門… 是的,就是當年這樣長時間的專注野外記錄,才有機會找到外觀相似的鈍頭蛇(Pareatidae,註)之間的細微差異。

從牧茂市郎在1931年發表的金絲蛇(Amphiesma miyajimae)和標蛇(Achalinus niger)之後,泰雅鈍頭蛇(Pareas atayal)是睽違84年以來再度在台灣發表的新種蛇類,同時也是首次由台灣人發表的新種蛇類!讓我們先暫停,倒帶一下,回到台灣第一次發現鈍頭蛇的年代吧。



1909年,美國兩爬學者范登堡(Van Denburgh)發表了台灣的第一種鈍頭蛇,正是廣為人知的台灣鈍頭蛇(Pareas formosensis)。稍後於1931年,日本兩棲爬蟲類大師牧茂市郎(Mouitirou Maki)於其鉅著《日本蛇類圖譜》中,發表了台灣所記錄的第二種鈍頭蛇,駒井氏鈍頭蛇(Pareas komaii)。
之後到了1997年,現代日本兩爬大師太田英利教授(Dr. Hidetoshi Ota)檢視了以上兩種鈍頭蛇的陳年標本外部形值,認為台灣鈍頭蛇和駒井氏鈍頭蛇事實上是同一個種(太田英利教授當年看的標本由於長時間以藥水浸泡保存,某些特徵例如虹膜顏色早已無法有所區別,再加上當年遺傳工具仍不是一個普遍的研究手段,因此可以理解為何太田教授當年會有這樣的結論)。根據命名規約,先發表先贏,駒井氏鈍頭蛇 (Pareas komaii)這個名字就被列為次同物異名,簡單來說就是被取消掉了。從此開始,台灣島上的鈍頭蛇又只剩下了一種。


後來的結果大家都知道了,在經由幾年瘋狂的野外攝影記錄,發現鈍頭蛇實際上很有可能有兩種甚至三種。帶著這個題目進到研究所,我的指導教授林思民老師費了很大的工夫把我教好(我應該超難教的XD),指導了我以往一竅不通的遺傳工具以及其它科學方法。然後我們終於把這個研究完成了!也在2015年的三月正式把泰雅鈍頭蛇發表在《動物文稿》(Zoologica Scripta)上,同時也恢復了駒井氏鈍頭蛇的地位。所以從這一刻開始,台灣的三種鈍頭蛇都已經被大家所接受、初步認識了。
接著疑問就來了:到底該如何區分這三種鈍頭蛇呢?首先最好區分的是台灣鈍頭蛇。台灣鈍頭蛇眼睛虹膜是橘紅色的 (圖7),體背部鱗片完全光滑無鱗脊 (圖8),頭部整體形狀也較短 (圖9)。



泰雅鈍頭蛇和駒井氏鈍頭蛇就不好分了。這兩種蛇的虹膜顏色都是黃色 (圖10),最主要的外觀差別在於鱗脊的強弱,而鈍頭蛇的鱗脊特別之處在於,是從體背部最中央的鱗列開始往兩側逐漸延伸、也逐漸減弱,直到完全消失變的光滑為止 (圖11)。


台灣的三種鈍頭蛇總鱗列數都是15列,然後我們如果說這條鈍頭蛇的鱗脊強,表示從牠體背部最中間那一列開始,往兩側延伸的列數較多;如果說鱗脊較弱,表示往兩側延伸的列數則較少。更簡單的來說,直接去數「帶有鱗脊的鱗列數」多寡就可以了。泰雅鈍頭蛇鱗脊較弱,根據我檢視過的標本,一般帶有鱗脊的鱗列數多在5列上下;而駒井氏鈍頭蛇的鱗脊較強,一般多在11列左右。


但生物不像工業製品,當然多少會存在個體差異,也因此仍然有些例外是難以按照這個通則來判定。所以若是可以搭配地理資訊來判斷的話,問題就會單純一些。如果這條黃眼睛的鈍頭蛇是在北部地區發現的(雪山山脈北側,大概是雙北、桃園、新竹、苗栗以及宜蘭),那就會是泰雅鈍頭蛇;除此之外在台灣中南部以及東部地區的黃眼睛鈍頭蛇,就是駒井氏鈍頭蛇了。

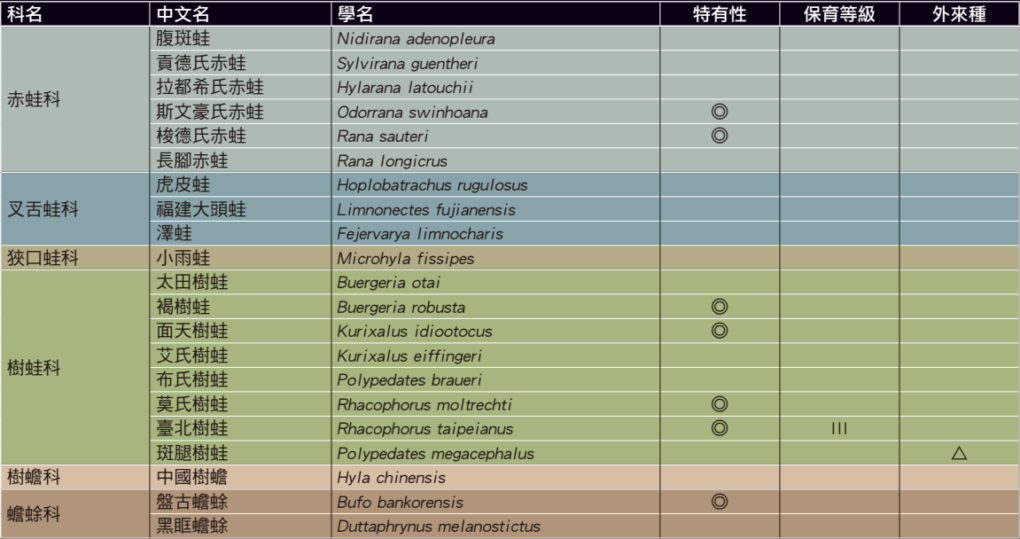
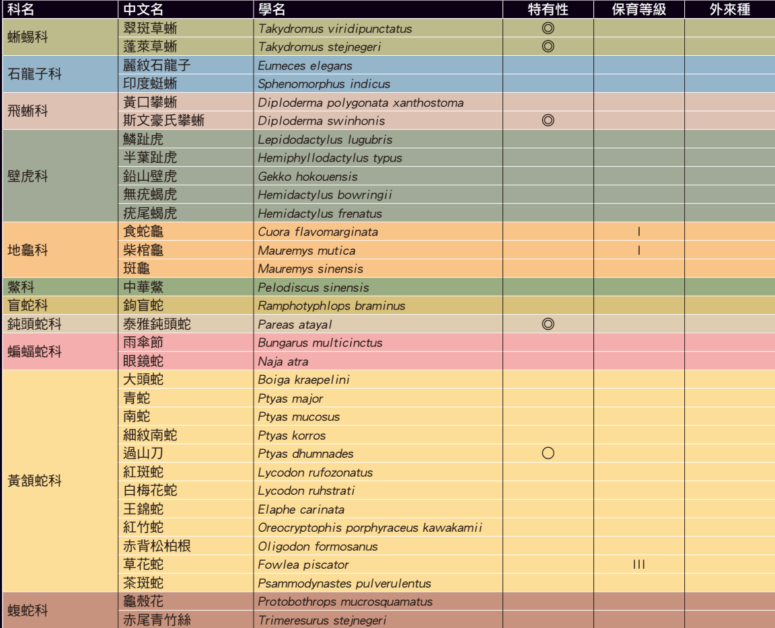
脊椎動物通常是人類最關注的一群動物,而一直以來蛇類更是人們相對害怕的動物。因為害怕,所以必須要更加的了解和研究。也因此在日治時代,可能是基於安全上的理由,嚴謹的日本科學家們幾乎已經將台灣蛇類資源調查完備了。所以84年後台灣人第一次發表本土特有蛇類:泰雅鈍頭蛇,無疑是一件非常難能可貴的事件。除了台灣陸生蛇類因此增加為48種(包含金門的緬甸蟒)、特有蛇類增為9種之外,我們更要多思考的是:「是否有某些生物在我們還不認識的時候就因為環境破壞而導致了滅絕?」
人類的生活一切資源都和生物多樣性息息相關。不管是食物、飲水,甚至藥物與科技,所有一切的原料與創意皆來自於自然,來自於生物多樣性。因此在這個氣候逐漸惡化、環境逐漸的不適合各類生物生存時,我們是否可以多一點的思考,少一點的消耗各種資源,繼續保護生物多樣性賴以依存的環境?保護生物多樣性就是保護人類的生存,希望藉由泰雅鈍頭蛇的發現,給大家提點一些雖然是老生常談但也不得不談的環保概念了。
註:
鈍頭蛇(Pareatidae)是一群體型小、側扁、樹棲的蝸牛蛞蝓專食者。只在熱帶以及亞熱帶亞洲地區分布的鈍頭蛇,是一群非常低調、溫馴且無害的小蛇。現有的鈍頭蛇科內包含了20個種,名錄如下:
- Aplopeltura boa
- Asthenodipsas laevis
- Asthenodipsas lasgalenensis
- Asthenodipsas malaccanus
- Asthenodipsas tropidonotus
- Asthenodipsas vertebralis
- Pareas atayal (泰雅鈍頭蛇)
- Pareas boulengeri
- Pareas carinatus
- Pareas chinensis
- Pareas formosensis (台灣鈍頭蛇)
- Pareas hamptoni
- Pareas iwasakii
- Pareas komaii (駒井氏鈍頭蛇)
- Pareas margaritophorus
- Pareas monticola
- Pareas nigriceps
- Pareas nuchalis
- Pareas stanleyi
- Pareas vindumi
而之所以近年鈍頭蛇在科學界的關注程度會變高,主因應該是京都大學細 将貴博士(Dr. Masaki Hoso)發現了鈍頭蛇的「慣用右手性」以及隱藏在背後的「與蝸牛共演化」等特性,是一系列非常了不起且衝擊學界認知的研究!

最後,感謝所有在研究期間給予協助的師長、朋友丶前輩和後輩,以及一條條因研究而犧牲的蛇蛇,更感謝台灣獨特的原住民族,泰雅鈍頭蛇因而借到了更多的光采,謝謝!
研究文獻: