本文由民視《科學再發現》贊助,泛科學獨立製作
目前人類已知的疾病散播途徑,可能要再增添一筆。2014年5月在PNAS上發表的一篇研究,指出了可能是人類史上第一個發現以風媒散播的發病因子,是甚麼病呢? 這是一種在1960年代在日本發現,但迄今仍未發現病源的川崎氏病。
甚麼是川崎氏病?
川崎氏病是一種好發於幼兒的疾病(年齡為六個月至五歲以下),這種病的流行期集中在每年的四月至六月,最初由小兒科教授川崎富作 (Tomisaku Kawasaki) 於1961年在日本首度紀錄;病發初期可注射抗體治療,若初期未治療妥當,則後期會發生冠狀動脈發炎甚至引起心臟病。這個疾病的成因至今仍是個謎,對於是否是單純因環境引起的自體免疫,還是由病原體引起的傳染性疾病,仍未有定論。
到底如何發病?
川崎氏病在固定月份引起高峰期的現象,吸引了理論生態學家羅多 (Xavier Rodó) 的注意,並帶領西班牙的研究團隊與日本合作開始對這現象展開研究。他們注意到日本川崎氏病的高峰期與西北氣流在時間上有重疊,這可能暗示了這種疾病的發生與兩國之間的空氣對流有關係,但要怎麼檢測它們有關聯呢?
他們首先使用了一種電腦模擬系統FLEXPART,這是一種藉由運算大氣流動方向來尋求其中的物質歷史軌跡的軟體,運算後發現,在四月到六月期間,日本受到大量西北風的吹襲,而這些風的路徑在地理上與大陸東北有高度重疊(圖一),在時間上,與日本40幾年來累積的疾病通報資料之川崎氏病高峰期也吻合;另外,調查後發現,大陸東北有著大量的農業區。
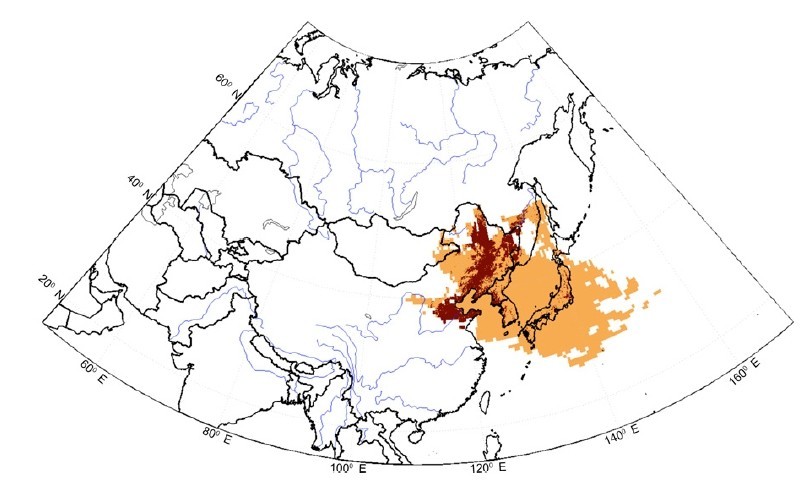
傳染還是非傳染?
為了釐清川崎氏病的流行,是否肇因人對人的傳染而引起? 研究團隊調查了東京及其他鄰近城市的發病情形,邏輯是,如果疾病是透過傳染而流行起來,則城市與城市之間的疫情,會有時間差,於是他們建立了一個簡易的易感-暴露-感染-復原模型(susceptible-exposed-infected-recovered, SEIR),輸入各城市的川崎氏病歷史回報資料,看是否有甚麼蛛絲馬跡可循,結論是,目前沒有已知的傳染病有這麼高的傳染效率,因為城市間的疾病通報資料,顯示時間差非常的短,即使把目前已知潛伏期最短的感冒病毒(如鼻病毒及B型流感),再縮短它的潛伏期十倍,在城市間散佈的時間仍高於目前觀察到的資料,因此排除了川崎氏病的流行是走人傳人的途徑。如果不是傳染,那是為什麼呢?很可能是所有的易感個體幾乎同時曝露在發病源下,這也與前者的風向說不謀而合。
風裡到底有甚麼?
為了瞭解這些風與川崎氏病的關係,研究團隊想出蒐集了空氣中的散布物資訊,看看裡面到底有甚麼,於是他們在東京市蒐集空氣中花粉及化學物,但這些物質都與川崎氏病的高峰時間不一致;接著,他們進行分析與川崎氏病流行期重疊的高空氣流內的微生物相(microbiota),他們在高度約2000~3000公尺的對流層(tropospheric)蒐集微生物,並與在地表附近蒐集的對照組看看有甚麼差異;這邊,是利用空氣中含有的微生物樣本下去做PCR擴增實驗,因為只要蒐集到生物,那就會有DNA,有了DNA並進行特定分類群片段的PCR擴增時,若蒐集到的生物裡面,有符合我們特定分類群片段的DNA,那就會有反應,如果沒有該分類群,那就不會有反應(因為沒有該分類群,就不會有該分類群特有的DNA片段,PCR對該片段的合成也不會成功)。
研究團隊就是利用細菌與真菌特有的16S與18S rRNA基因片段進行PCR擴增後,發現了在高空的樣本中充滿大量念珠菌屬(Candida sp.)物種的反應,而地表附近的樣本在經過PCR後只發現麴黴屬(Aspergillus sp.)的反應(圖二)。目前觀察發現,真菌與農作物有相當常見的共生或寄生關係,而念珠菌在動物實驗上,也的確展示出部分與川崎氏病類似的症狀,因此在綜合了檢測大氣中的微生物、測量大氣對流的源頭並且排除傳染的可能性後,在日本的川崎氏病,也許與經由大氣散播的的念珠菌或其製造的毒素有關。

後記
本研究揭露了在日本的川崎氏病,其發病原因與高空對流帶來的念珠菌有潛在的關聯性,但並沒有辦法直接推論是念珠菌引起的,畢竟這些念珠菌是不是真的由中國的農業區來的還不知道(這可能要再跟中國合作調查農作物),也沒有明確指出念珠菌產生了甚麼東西讓日本的小孩發病,因為目前已知的真菌毒素(mycotoxin)中,並沒有任何一種會讓人類引起川崎氏病的症候群;再來是,川崎氏病並非日本獨有的疾病,在美國本土也有發生,那有任何類似的大氣對流活動也發生在美國嗎? 或進一步問,美國的川崎氏病是不是也是因為這種大氣對流帶來的毒物引發的呢? 這些都有待解釋,但這篇研究的貢獻在於,他們用比較巨觀的角度去看一個疾病發生的原因,結合生態學觀點、數學模型與歷年醫學資訊去探索川崎氏病這塊雜亂的拼圖,我認為是非常好的一個起點,相信未來對美國川崎氏病的研究會帶來更多的亮點,為這塊雜亂的未知圖像補上更完整的場景。
延伸閱讀:
- 川崎氏病-Wikipedia
- FLEXPART的網站,內有文獻可得知運算機制。
- SEIR只是疾病擴散的其中一種模型,可至下方wiki連結見到各種擴散模型介紹。
- PCR,聚合酶連鎖反應(Polymerase Chain Reaction),是目前生物科學廣泛使用的工具,原理可見這個很直白的卡通影片。
原始文獻:
Rodó, et al. 2014. Tropospheric winds from northeastern China carry the etiologic agent of Kawasaki disease from its source to Japan. Proceedings of the National Academy of Sciences, early edition. doi: 10.1073/pnas.1400380111
—————————–
更多內容也可以上科技大觀園搜尋「細菌」,或每週六上午8點收看民視53台科學再發現。