- 文/黃尹則|佛羅里達大學博士,現任京都大學博士後研究員。喜歡從共生學的角度來看生物。貓奴。
大約一萬兩千年前,世界各地的人類不約而同地從狩獵、採集的生活型態轉型成遊牧、群聚的農業聚落。農業行為的出現,深深地影響了人類發展的腳步;人口的急遽增長、專業分工的產生(有更多時間去做找食物以外的事情),間接導致社會結構的出現,爾後組成大型聚落甚至是政府。
因此,發展出農業行為是人類演進過程中的重大事件。
但相較於其他生物,其實人類在這場「農業遊戲」中是很晚才加入的玩家。
約一億年前,一群住在樹皮以下、身形不到一公分的小甲蟲就已經開始種植他們的真菌作物。這類甲蟲稱為菌蠹蟲 (Ambrosia beetle),而其真菌作物稱為菌蠹蟲真菌 (Ambrosia fungus)。
菌蠹蟲和菌蠹蟲真菌之間是一種絕對 (obligate)、互利 (mutualistic) 的共生關係。真菌是菌蠹蟲的唯一食物來源,在沒有真菌的情況下,菌蠹蟲無法存活。而菌蠹蟲是這些真菌在環境中傳播的唯一交通工具,自然界中,菌蠹蟲真菌只存在於菌蠹蟲的棲息環境中。兩者相依相存、缺一不可。

絕大部分的菌蠹蟲棲息於新鮮剛死亡或是極度衰弱的的樹木,只有極少數的種類會攻擊健康的樹木(例如造成美國酪梨產業嚴重危害的幾種菌蠹蟲)。
樹木在衰弱時,它們會釋出酒精及其他化學物質,菌蠹蟲就是藉由這些揮發物質來選擇合適的樹木寄主。當先驅的菌蠹蟲飛到合適的樹木上,找到自己喜歡的位置後,便開始用其強壯的下顎 (mandible) 和全身的力量突破樹皮,深入到木質部挖掘出屬於自己的家。
菌蠹蟲的家因為長得很像「隧道」,英文便直接用 gallery 來描述菌蠹蟲的棲所。在挖掘隧道的同時,菌蠹蟲一邊產卵、一邊將共生真菌接種在隧道壁上,當卵孵化時,共生真菌已培養成豐盛的真菌花園 (fungal garden),以提供幼蟲生長發育時所需營養。
當幼蟲成長、化蛹、再蛻變成新一代的成蟲後,便攜帶其共生真菌,離開隧道去尋找下個合適的樹木寄主。單一世代的生活史大約只需要一到三個月的時間。

如同我們所熟悉的人類農業,菌蠹蟲與共生真菌的演化過程中也包含了「馴化」、「性狀管理」、和「病害控制」等複雜行為。
農業的起點:馴化事件
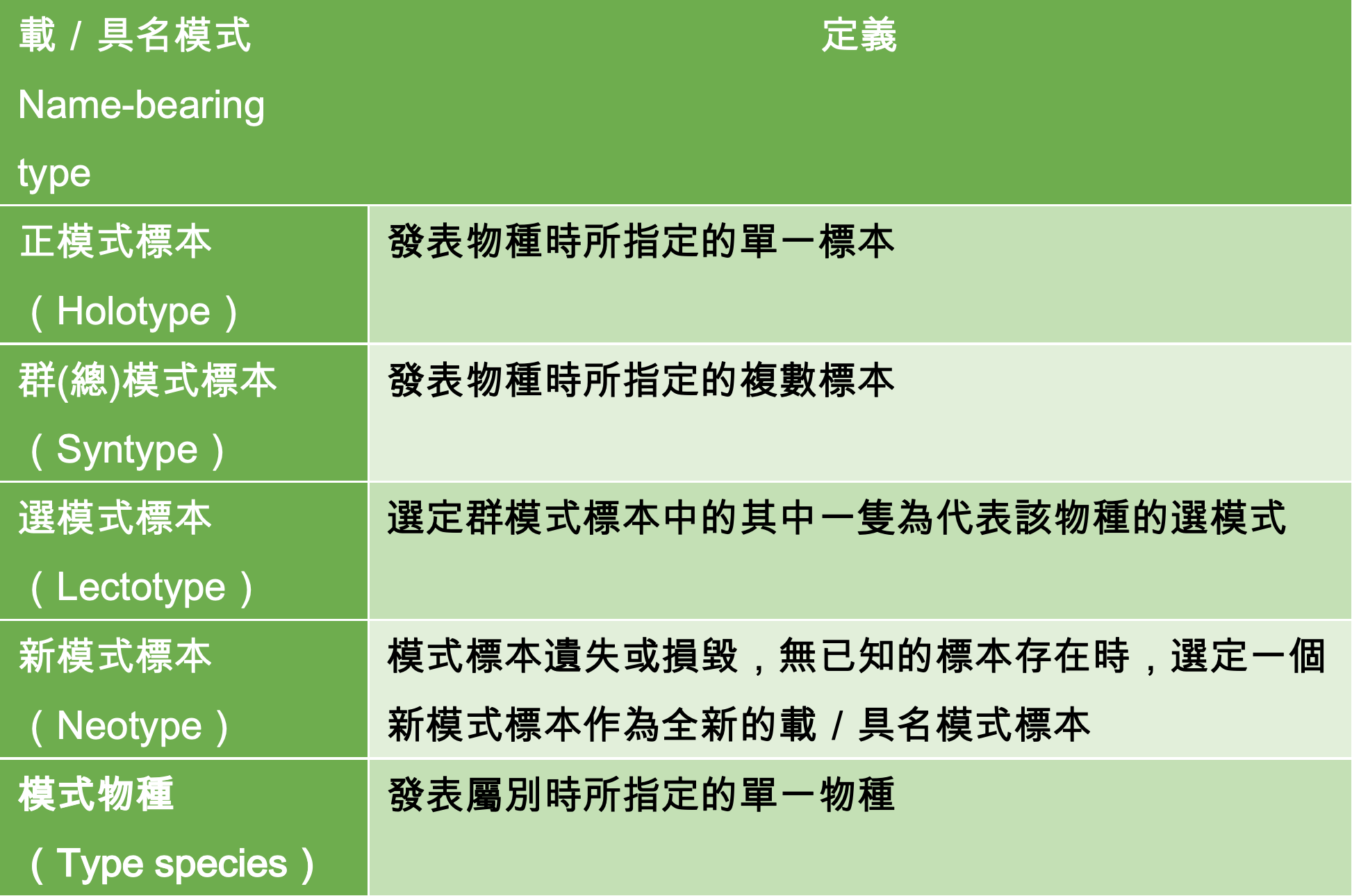
雖然至今仍無法窺探菌蠹蟲和共生真菌實際的馴化過程,科學家還是能透過基因親緣樹來推斷演化歷史上共發生過幾次「馴化事件」。現今已知的菌蠹蟲約 3400 種,而菌蠹蟲真菌則有 3000 餘種。
事實上,菌蠹蟲並非單一種昆蟲,他們泛指小蠹蟲亞科 (Scolytinae) 和長蠹蟲亞科 (Platypodinae) 內具有農業行為的多個類群。根據小蠹蟲的親緣樹推斷,其演化過程中至少有 12 至 16 次的演化支系 (lineage) 獨立演化 (indepedently evolved) 出農業行為。同樣的,菌蠹蟲真菌也不是單一種真菌,而是一群與菌蠹蟲形成共生關係的真菌。在真菌的親緣樹上,目前已知至少發生了 13 次的馴化事件。
看到這裡,聰明的你有發現農夫(菌蠹蟲)與作物(菌蠹蟲真菌)間演化次數的不同嗎?沒錯,菌蠹蟲與共生真菌並不完全是一對一的關係。
有些菌蠹蟲只攜帶一種真菌、有些則攜帶多種真菌;有些菌蠹蟲真菌只被一種菌蠹蟲豢養、有些則會出現在不同種的菌蠹蟲的隧道裡。但神奇的是,無論是一對一或者一對多的關係,這樣的共生關係在多數環境中是穩定的(維持一對一或一對多)。他們是如何做到的呢?這就要提到他們精緻的管理模式。
性狀管理
在長達一億年共同演化的過程中,菌蠹蟲和真菌為了彼此的共生關係,各自發展出了精巧的構造。菌蠹蟲為了有效攜帶共生真菌的孢子,在外骨骼上或是在體內「挖」出了一種特殊構造──儲菌器 (mycangium)──來確保每一世代的成蟲所攜帶的真菌都是同一種類,而非環境中其他種隨機的真菌。
科學家透過實驗證明,發現某些儲菌器具有「選擇」正確共生真菌的能力,而某些則具有比較大的彈性能容忍「相近但不同種」菌蠹蟲真菌入住的現象。

而從真菌的角度來看,首先要先知道,作為傳播用途的真菌孢子體型一般不會太大;裡面所含的養分僅需足夠讓其發芽、長出菌絲,直到找到新的營養來源即可。部份菌蠹蟲真菌在數千萬年的演化過程中,被選擇出能產生不成比例的巨大孢子類型。這些巨大孢子有較多的空間來儲存醣類、脂肪、胺基酸等昆蟲成長所需的營養。
但目前在性狀管理的機制上,例如儲菌器如何維持專一性的共生關係(物理性或化學性?)、菌蠹蟲真菌的營養性狀是如何被選擇的,是目前科學家們尚未釐清的問題。
病害控制:社會性分工與世代交替
與人類的農業相似,菌蠹蟲與其真菌花園也受到許多潛在病害的威脅。這些「病原」可能來自於先驅菌蠹蟲體表上的伺機性微生物(例如非共生真菌、細菌、線蟲、蟎等),還有會獵食菌蠹蟲的天敵昆蟲等。
那菌蠹蟲如何去對抗這些威脅呢?前面有提到,人類因為農業行為的產生,進而發展出複雜的社會結構。同樣的,部份種類的菌蠹蟲也具有社會性特徵:有世代重疊的現象(族群內同時有不同階段的個體)、共同育幼的行為、分工制度。
科學家發現,某些菌蠹蟲為了維持隧道內的清潔,幼蟲負責將排遺滾成球狀 (balling)、成蟲將隧道中的木屑、死亡的個體推至洞口 (shuffling)、最後由負責堵住洞口的成蟲將廢棄物排出。同時,這些負責堵在洞口的成蟲肩負了抵擋天敵入侵的責任。此外,當族群內大多數的成蟲離巢或死亡後,其真菌花園很快就會被其他伺機性真菌及細菌所污染並且取代,進而導致整個隧道內族群的瓦解。
因此,即使菌蠹蟲單一世代的生活史很短(大多僅一至三個月),仍需要家族中所有成員的分工合作才能確保整個族群的延續。
了解菌蠹蟲與真菌共生特性的重要性在哪?
菌蠹蟲與菌蠹蟲真菌的共生關係,不管從生物學的角度或者是應用方面都有很高的研究價值。
生物學方面,由於演化過程中它們多次獨立演化出農業行為,透過比較這些獨立發生的事件,可以幫助我們釐清昆蟲與真菌共生關係是如何開始的。
應用方面,雖然攻擊健康樹木的菌蠹蟲種類很少,但幾乎都造成生態系等級的危害。例如前面提到危害美國酪梨產業的菌蠹蟲,其中一種就是從台灣被引進美國的入侵種。了解菌蠹蟲和共生真菌的生態,將有效幫助我們控制他們所造成的危害。
引用文獻
- Johnson AJ, McKenna DD, Jordal BH, Cognato AI, Smith SM, Lemmon AR, et al. Phylogenomics clarifies repeated evolutionary origins of inbreeding and fungus farming in bark beetles (Curculionidae, Scolytinae). Mol Phylogenet Evol 2018; 127: 229–238.
- Vanderpool D, Bracewell RR, McCutcheon JP. Know your farmer: Ancient origins and multiple independent domestications of ambrosia beetle fungal cultivars. Mol Ecol 2018; 27: 2077–2094.
- Hulcr J, Stelinski LL. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu Rev Entomol 2017; 62: 285–303.
- Skelton J, Johnson AJ, Jusino MA, Bateman CC, Li Y, Hulcr J. A selective fungal transport organ (mycangium) maintains coarse phylogenetic congruence between fungus-farming ambrosia beetles and their symbionts. Proceedings of the Royal Society B: Biological Sciences 2019; 286: 20182127.
- Huang Y-T, Skelton J, Hulcr J. Lipids and small metabolites provisioned by ambrosia fungi to symbiotic beetles are phylogeny-dependent, not convergent. ISME J 2020; 14: 1089–1099.