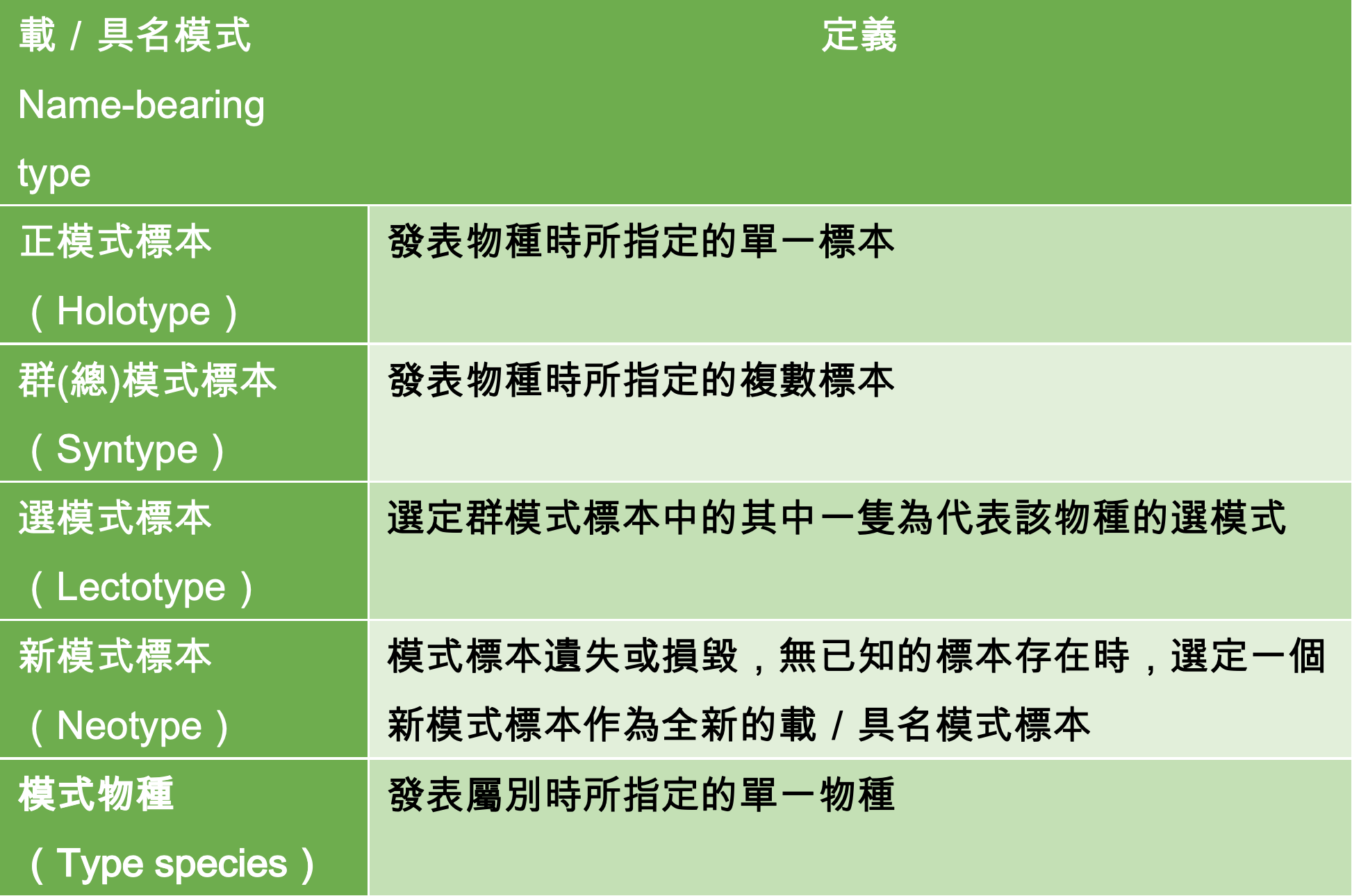
編按:本文為台灣魚類學家何宣慶自述近期發表的擬棘茄魚屬(Halieutopsis)分類研究背後的故事。
這是一個跨越兩個世代研究一個魚類類群的故事,事情開始於 17 年前的一個新種以及我的一趟美國之旅。
研究船帶回來的新種
在 2005 年一月,台灣的深海調查仍如火如荼在進行中,當時我剛剛結束兵役,回到中研院的實驗室。助理帶回來一些棘茄魚的樣本,希望我可以鑑定種類。第一隻樣本是一隻灰灰的,全身長滿小刺的小魚,全長只有四公分左右,大概可以知道是擬棘茄魚屬(Halieutopsis),但是翻遍文獻完全無法得知是什麼種。

為了進一步破解這個問題,我們寫信問了當時已經自丹佛大學退休的 Dr. Margaret G. Bradbury,她從 1960 年代開始研究這個類群,發表過不少重要的相關著作。她告訴我們,她想起了在夏威夷的主教博物館(Bishop Museum)也有看過類似的樣本,建議我們可以借回來進行比對看看。很快地我們就確認這個物種是新種,並以 Dr. Bradbury 的名字命名為瑪格莉特擬棘茄魚(Halieutopsis margaretae Ho & Shao, 2007)。
擬棘茄魚屬是一種小型的鮟鱇魚,它們的身體相當扁平,體盤接近圓盤狀或略為心形,具有一細細的尾部,看起來有點像是很小的魟魚。它們隸屬於鮟鱇魚目下棘茄魚科的成員,所有種類都是深海魚,最深的紀錄達四千公尺以下,除了是棘茄魚最深的紀錄外,也是鮟鱇魚中最罕見的一群。一般研究調查鮮少有機會採集到樣本,而少部分物種即使樣本數較多的樣本,也都來自於是研究船在人煙罕至的地方所採集而來的,平時的漁港很難找到,如台灣的樣本多數也是來自靠研究船的調查而來。


跟上一個世代的棘茄魚分類權威交流
2005 年 5 月,我獨自前往美國幾個研究單位進行研究,並在美國的魚類與爬蟲類學會年會上報告了新種的擬棘茄魚。因為還尚未確定博士班題目,所以還算是在摸索的階段。在加州科學院停留的時候,聽聞 Dr. Bradbury 就住在南部靠近 Monterey Bay 的 Pacific Groove,於是我就跟她約定時間,並央請一位博士生載我去拜訪。這次的拜訪,我們稍微交換了一下研究的經歷,多數是我聽她說,我提出一些問題,她也很認真回答。拜訪中,她很快拿出一疊又一疊的底片、黑白照片還有許多寫了密密麻麻的筆記跟不少的文獻,並說這些都給我:「Good luck! Young man !」隨後我們一起簡單用過午餐後就道別。
接下這些資料之後,當下除了感動也覺得有些責任跟義務,需要好好地做研究。這也就促使我選定「棘茄魚的分類及系統演化研究」當成博士研究的主題。隔一年,我又再次回到加州科學院,也再次拜訪 Dr. Bradbury,將我一年的收穫跟她說明,她顯得非常開心,一直都笑著聽我說,中途也會很直接地告訴我她的想法。
印象很深刻的是,其中有一隻樣本與我的鑑定不同,我問她為什麼會把這個魚鑑定成這個物種?她只是笑笑告訴我:「其實我也不太確定,但是因為他們需要放一個名字,所以我就給他們一個名字……。」當下其實覺得她很豁達,這也讓我比較放心,把那個樣本正確的學名寫上去。當然除此之外,她還有給我更多她做棘茄魚分類的邏輯跟緣由,這讓我可以追溯她過去所做的研究以及思考如何做判斷。
Dr. Bradbury 先前的研究
時序回到 1988 年,Dr. Bradbury 已經發表了全球的擬棘茄魚分類研究,並描述四個新種,在依靠郵件通信跟船運的年代,要獲得這些樣本並進行研究已經是相當不易,尤其是有些樣本甚至於來自丹麥哥本哈根、俄國莫斯科等地的館藏。往後的數十年間,Dr. Bradbury 並沒有停止對於這個類群的研究,而是親自前往俄國、印度、歐洲各個不同的館所檢視樣本、收集資料,還有用她高超的攝影技巧拍攝許多精美的標本照。這一批珍貴的資料,也都在她先前所贈與我的資料中。
有了這樣的基礎後,我除了在不同的國家跟博物館間穿梭,檢視樣本外,也按圖索驥,回到過去 Dr. Bradbury 去過的地方,實際檢視樣本,逐一確認。包含當時令她覺得困惑的近似種。
站在巨人的肩膀上,跨越兩個世代的研究結果
分類研究包含了一連串不斷的考證,可能也是某種形式的考古或文獻探討。上一世代留下未解的謎題與資料,讓後面的人可以繼續進行、進一步釐清相關的疑惑。有幸可以在承襲 Dr. Bradbury 的研究下,讓我可以站在巨人的肩膀上看世界。
因為擬棘茄魚屬跟腔蝠魚屬(Coelophrys)及雙鰓魚屬(Dibranchus)相當接近,所以中間有不少的混亂或誤鑑的狀況。所幸在有博物館跟研究計畫持續地支持下,讓我得以持續進行研究。歷經將近十七年的時光,也才終於把所有的資料都準備完備,並且撰寫成論文發表。這大概也有賴於疫情期間,無法出國去做研究,所以就坐下來好好整理資料,並且投稿發表。
解謎擬棘茄魚屬:前人留下的謎題以及物種釐清
有幸於在 2021 年發表的這個文章,主要釐清了過去所發表的所有學名的有效性以及討論同種異名。擬棘茄魚屬過去還存留有不少有疑問的物種,這也困擾著幾個世代的人們。
首先是由英國著名的自然科學家 A. W. Alcock 在 1891 年所發表的 Dibranchus nasutus。Alcock 最著名的是在印度的 Zoological Survey 工作時所發表的許多魚類巨作,其中包含許多精美的繪圖。不過,等到 Dr. Bradbury 在 90 年代見到 D. nasutus 的正模標本時候,只有在瓶中看到魚體的碎片,而且還包含兩個個體。我個人在 2015 年造訪時也再次印證這件事。而且,當時Alcock並未出版這一個物種的繪圖。因此,在 Alock 發表後的往後幾十年間,沒有人能確定 D. nasutus 這個物種的歸屬。
從標本碎片中,Dr. Bradbury 已經約略可以看出這個種很接近擬棘茄魚,但不確定是哪個物種。而我在檢視全球的樣本之後,發現原來 Alcock 曾經寄送一尾由他親自鑑定的標本到大英自然史博物館,這個發現讓我可以很快確認,它跟後來在菲律賓所發表的蟲紋擬棘茄魚(Halieutopsis vermicularis)是同一個物種。因此在這次的發表中,重新將 D. nasutus 歸入擬棘茄魚屬成為大吻擬棘茄魚(Halieutopsis nusuta),並將蟲紋擬棘茄魚歸入其次同物異名(junior synonym)。

第二個有問題的物種,則是同樣印度進行研究的英國學者 R. E. Lloyd 在 1909 年利用緬甸外海採集的樣本所發表的 Dibranchus nudiventer,這個物種的特徵是蠻明顯的,只是模式標本已經遺失,無從考究。所以先前 Dr. Bradbury 在幾個文獻中一開始以疑似蟲紋擬棘茄魚(Halieutopsis cf. vermicularis)稱之,後以棘茄魚科中的未定種(Incertae sedis)稱之。這也間接導致日本學者(Shimazaki et al. 2004)錯誤地將日本的樣本誤認為是真正的蟲紋擬棘茄魚。
事實是從原始的描述跟其他文獻中,可以知道蟲紋擬棘茄魚的尾部棘為雙叉,D. nudiventer 的尾棘為單一不分叉。我也發現,在 Dr. Bradbury 許多的標本照中,有一張在莫斯科大學拍的,她已經觀察到,有另一個長得很像是她所發表的 Halieutopsis bathyoreos 的物種,所以將兩者放在一起做比較並拍照,而這一個物種正是真正的 D. nudiventer。經過檢視許多印度洋的樣本,並且多方以各種文獻做比較,可以確認這是擬棘茄魚屬中的一個有效種,因此更名為裸腹擬棘茄魚(Halieutopsis nudiventer)。

第三個有問題的物種是 Coelophrys oblonga,這個物種的模式標本產自菲律賓,唯一個正模標本曾經乾掉過,所以現在已經是嚴重縮水的狀態,除了一些身體表面的棘還可以清楚辨識以外,很多特徵都無法確認。Dr. Bradbury 認為這可能是雙鰓魚屬的一種,但不確定是哪一個物種。首先因為腔蝠魚屬的腹鰭很小,而這個物種有正常大小的腹鰭,所以應不屬於這個屬。一開始我們在水試所的樣本中發現一尾採自南中國海的樣本,這一個樣本吻部有相當特殊的棘刺,我們一度認為是新種。不過在比對過橢圓腔蝠魚的模式標本後,確認兩者應該是一樣的,因此我們將這個物種歸入擬棘茄魚屬中,成為橢圓擬棘茄魚(Halieutopsis oblonga)。

為什麼有這麼多有問題的物種
其實先前擬棘茄魚屬這個類群中被誤鑑機率最高的就是大吻擬棘茄魚,不過可能主要是因為這是最早被描述的物種。
舉例來說,Brauer(1906)年曾經提到這個種,但是我們實際檢視其描述內容才發現,其實是後來 1988 年發表的 Halieutopsis bathyoreos 的誤鑑。而 Norman(1939)在報導 John Murray Expedition 的採集結果時,也將一尾採自紅海的樣本鑑定為大吻擬棘茄魚,我實際檢視這尾保存在大英自然史博物館的標本時,發現它的各種棘刺樣式跟形態都跟大吻擬棘茄魚不同,也跟其他同屬成員不同。於是在歷經近一個世紀後,才得以被正式描述及命名,我將它命名為新種莫氏擬棘茄魚(Halieutopsis murrayi)。另外一個被認定為大吻擬棘茄魚的則出現在 Radcliffe(1912)所發表的文獻中,但是完全沒有機會進行驗證,因為當時只有一張圖片,標本已經無從考證,因此只能留待以後有機會再次在菲律賓採集到時才能做確認。日本學者所認定的蟲紋擬棘茄魚(現為大吻擬棘茄魚之次同物異名),現在則被我重新鑑定為裸腹擬棘茄魚。
當然,會有這麼多問題存在主要還是在於標本過於稀少,能夠全面研究的機會不多。再者,可以參考的文獻也不多,所以這個類群一直處於不明的狀態,一直到今日才被完整研究。
重新分類後的成果
最後發表的研究共計彙整了全球 16 個擬棘茄物種資訊,包含發表 5 個世界新種,其中有 3 個新種是來自台灣。分別是:
- 來自台灣東北海域所採集的新種台灣擬棘茄魚(Halieutopsis taiwanea)。

- 採自台灣及澳洲昆士蘭的棘皮擬棘茄魚(Halieutopsis echinoderma)。

- 採自台灣及印尼,以日本學者命名的河合氏擬棘茄魚(Halieutopsis kawaii)。

- 採自紅海的莫氏擬棘茄魚。

- 以日本學者岡村收(Osamu Okamura)之名,所命名的岡村氏擬棘茄魚(Halitueopsis okamurai)。

橫跨半世紀的經驗傳承將持續下去
擬棘茄魚屬這個研究,所涵蓋的物種命名與採集跨越了 130 年,也歷經了兩個世代(1960 年代至今)的資料累績才得以完成。
從我與 Dr. Bradbury 第一次見面交換心得,到後面歷經所有到不同博物館做研究的經驗、體驗不同國家的民俗風情、入住不同的青年旅館、在不同時區穿梭於各個機場、以及搭乘不同的交通工具,許多情景都歷歷在目。
或許幾十年的時間並不長,但是能夠持續不斷傳承,並得以完成算是相當不容易。我曾邀請 Dr. Bradbury 一起發表這篇論文,但她相當客氣地婉拒,認為年輕人自有一片天。遺憾的是 Dr. Bradbury 在 2010 年我完成博士學業之後不久就過世,無法親眼見到這篇研究的發表。
僅以本篇文章紀念 Dr. Bradbury 對我的研究影響以及對學界的貢獻。
- 相關分類學研究發表於 2021 年 12 月 30 日刊登於 Journal of Marine Science and Engineering。
詳見:Ho, H.-C. 2021. Taxonomy and distribution of the deep-sea batfish genus Halieutopsis (Teleostei: Ogcocephalidae), with descriptions of five new species. Journal of Marine Science and Engineering, 10(1): 1-64.
參考文獻
- Bradbury, M. G. 1988. Rare fishes of the deep-sea genus Halieutopsis: a review with descriptions of four new species (Lophiiformes: Ogcocephalidae). Fieldiana Zoology (New Series), 44: 1-22.
- Ho, H.-C. and K.-T. Shao. 2007. A new species of Halieutopsis (Lophiiformes: Ogcocephalidae) from western North and eastern central Pacific Ocean. Raffles Bulletin of Zoology Suppl. no. 14: 87-92.
- Shimazaki, M., H. Endo and M. Yabe. 2004. Redescription of a rare deep-sea batfish, Halieutopsis bathyoreos (Lophiiformes: Ogcocephalidae). Ichthyological Research v. 51(no. 2): 120-125.