參考文獻:Hsiao, Y (2015) Description of two new species of the genus Stenothemus from Taiwan (Coleoptera: Cantharidae). Zootaxa 3937: 386–392. doi: 10.11646/zootaxa.3937.2.9
1990 年,融合蛋白 CD4 免疫黏附素(CD4 immunoadhesin)誕生。這項設計,是為了對付令人類聞風喪膽的 HIV 病毒。
-----廣告,請繼續往下閱讀-----
我們知道 T 細胞是人體中一種非常重要的白血球。在這些 T 細胞中,大約有六到七成表面帶有一個叫做「CD4」的輔助受體。CD4 會和另一個受體 TCR 一起合作,幫助 T 細胞辨識其他細胞表面的抗原片段,等於是 T 細胞用來辨認壞人的「探測器」。表面擁有 CD4 受體的淋巴球,就稱為 CD4 淋巴球。
麻煩的來了。 HIV 病毒反將一軍,竟然把 T 細胞的 CD4 探測器,當成了自己辨識獵物的「標記」。沒錯,對 HIV 病毒來說,免疫細胞就是它的獵物。HIV 的表面有一種叫做 gp120 的蛋白,會主動去抓住 T 細胞上的 CD4 受體。
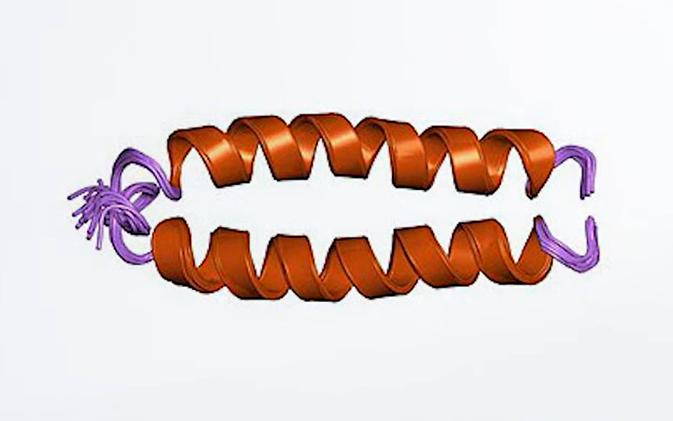
而另一端的 Fc 區域則有兩個重要作用:一是延長融合蛋白在體內的存活時間;二是理論上能掛上「這裡有敵人!」的標籤,這種機制稱為抗體依賴性細胞毒殺(ADCC)或免疫吞噬作用(ADCP)。當免疫細胞的 Fc 受體與 Fc 區域結合,就能促使免疫細胞清除被黏住的病毒顆粒。
不過,這裡有個關鍵細節。
在實際設計中,CD4免疫黏附素的 Fc 片段通常會關閉「吸引免疫細胞」的這個技能。原因是:HIV 專門攻擊的就是免疫細胞本身,許多病毒甚至已經藏在 CD4 細胞裡。若 Fc 區域過於活躍,反而可能引發強烈的發炎反應,甚至讓免疫系統錯把帶有病毒碎片的健康細胞也一併攻擊,這樣副作用太大。因此,CD4 免疫黏附素的 Fc 區域會加入特定突變,讓它只保留延長藥物壽命的功能,而不會與淋巴球的 Fc 受體結合,以避免誘發免疫反應。
從 DNA 藍圖到生物積木:融合蛋白的設計巧思
融合蛋白雖然潛力強大,但要製造出來可一點都不簡單。它並不是用膠水把兩段蛋白質黏在一起就好。「融合」這件事,得從最根本的設計圖,也就是 DNA 序列就開始規劃。
我們體內的大部分蛋白質,都是細胞照著 DNA 上的指令一步步合成的。所以,如果科學家想把蛋白 A 和蛋白 B 接在一起,就得先把這兩段基因找出來,然後再「拼」成一段新的 DNA。
而若提到長得不像隱翅蟲的隱翅蟲,那也不得不提四眼隱翅蟲亞科(Omaliinae)了。這個亞科的隱翅蟲有一部份是長相怪異的傢伙,像是分佈在臺灣高海拔山區的蔣氏四眼隱翅蟲(Deinopteroloma chiangi)就是其中一個例子,牠的特色就是他的翅鞘已經完全包覆住腹部了。這個屬的隱翅蟲原來發表時被當作是埋葬蟲科(Silphidae)的一員,但 1985 年由著名的隱翅蟲分類學家 Dr. Ales Smetana的形態分析結果,支持該屬為隱翅蟲科,且應被放置在四眼隱翅蟲亞科中(Smetana, 1985)。
Jaloszynski, P. 2018. World genera of Mastigitae: review of morphological structures and new ecological data (Coleoptera: Staphylinidae: Scydmaeninae). Zootaxa 4453 (1): 001-119.
Grebennikov V. V. & Newton, A. F. 2009. Good-bye Scydmaenidae, or why the ant-like stone beetles should become megadiverse Staphylinidae sensu latissimo (Coleoptera). Eur. J. Entomol. 106: 275-301.
Smetana, A. 1985. Systematic position and review of Deinopteroloma Jansson, 1946, with descriptions of four new species (Coleoptera, Silphidae and Staphylinidae (Omaliinae)). Systematic Entomology 10 (4): 471-499.