

說人類是一種被下半身支配的奇怪動物一點也不為過─我們的確是為數不多會透過性愛取樂,非單純為了繁殖而交配的物種之一。然而,對於某些動物而言,性的過程或許和「愉悅」這的詞相距甚遠。早先發表的兩篇論文告訴我們,在某些情形中,性更像是一場戰役。從「小雞雞擊劍」到「愛之飛鏢」,這兩篇文章提到的性形式並無美妙的體位,反而更像重口味的SM方式。
1897年,義大利動物學家Constantino Ribaga在雌性臭蟲(Cimex lectularius)腹部的中上區發現了一個奇怪的器官。Ribaga最初猜測這是一個類似於蟬發聲器的結構。但是當他進行解剖時卻發現,在這個器官的腹側細胞數中存放著大量的精子。(這個器官後來被稱做受精儲精器spermalege)
這些精子是如何到達那裡的呢?當時的科學家們對這個問題束手無策。在他們的猜想中,雄性臭蟲用大量的精子潮水般地淹向雌性臭蟲(我的愛如潮水…),雌性臭蟲透過這個器官消化多餘的精子,就像接受聘禮一樣。但這個理論並不可靠…
直到1913年,人們才觀察到雄性臭蟲是透過一個恐怖的、注射器針頭般的生殖器「刺穿」雌性臭蟲的這個器官,並在這個創口上與雌蟲進行交配。精子通過體腔徑直地游向卵巢,這種交配方式被稱為「創傷性授精」。

本文介紹的第一篇文獻來自《生物學回顧》(Biological Reviews)。Rolanda Lange和來自德國杜賓根大學和英國謝菲爾德大學的同事共同發現,在很多無脊椎動物中都存在類似的破壞「性」活動。
作為雌雄同體生物,蝸牛有一種稱為「創傷性分泌物轉移」的性挑逗方式,透過近距離向潛在的交配對象發射一種被刺激神經的黏液包裹的「愛之飛鏢」(gypsobelum)來達成。可想而知,在這種求愛形式下沒有蝸牛希望被暗器射中─沒有人想要當M(好啦,對蝸牛來說就是不想要當女生)。某些情況下,被愛之飛鏢督中的一方在接受精子的同時也會反將一軍,再督回去正在督他的那個人(就是環形接龍的感覺拉)在另一種雌雄同體的生物扁蟲中,這種暴力性活動則以「陰莖擊劍」的形式呈現,雙方都試圖用陰莖捅傷對方,勝者透過創口強行授精。


為什麼一個雄性生物要如此殘暴地刺傷未來孩子他媽呢?今年1月發表在《昆蟲學年度回顧》(Annual Review of Entomology)的一篇文獻中,西澳大利亞博物館的Nik Tatarnic和同事們對節肢動物進行的深入研究。從演化的角度來看,他們把這種暴力行為解釋為雄性生物「改變局勢的戰術」。
要產生後代,交配當然是必不可少的過程,但是交配卻只是一個前奏。更加重要的是受精過程,而雌性生物顯然更希望控制受精的時間和地點還有孩子他爸是誰。在許多情況下,雌性在這件事上都做得非常成功,例如利用生殖道篩選出她比較偏愛的「客兄」。某些雌性動物能夠直接噴出或化學性過濾掉不受青睞的精子(這個現象稱為隱蔽雌性選擇 cryptic female choice),有時還可以完全關閉生殖器。雌性的受精調控在昆蟲中尤其普遍,雌蟲可以把精子存放在一個囊中,有時甚至可以存放好幾年,某天閒閒的時候拿出來一些來受精。
另一方面,對雄性而言,起初他沒考慮這麼多,總之行有餘力就多播幾個種,簽越多中獎機率越大。但你想一想,如果你的素質就是沒有高富帥,那不就跟「左手只是輔助」一樣嗎。所以有些動物發揮了一點創意,為了確保雌性所受精的都是自己的精子,雄性不但要戰勝雌性的受精防禦系統,還要擊敗雌性的其他配偶。這或許就是一場沒有結局的「兩性軍備競賽」。
雄性擊敗對手的第一步,就是靠著花枝招展的求愛表演來給雌性留下好印象,這或許將幫助他們在受精階段時拔得頭籌。但是這樣的競爭方式實在是過於文明,雄性常常是更加卑劣的─他們演化出一些奇葩的行為來確保自己的精子可以獲勝,例如在交配後封閉雌性生殖道來阻止其他人再亂督,或是直接舀出前幾個對手的精液。我們雄性人類的生殖器就有人推斷具有「精液挖勺」般的第二功能。
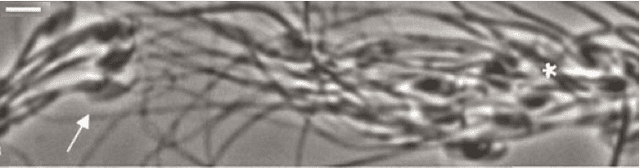
另一種辦法是正面強攻雌性的受精防禦系統。雄性果蠅在精子細胞中「加料」,利用化學物質促使雌性果蠅增加排卵量,儘管這個行為會導致雌果蠅免疫系統折損、壽命縮短。雌性小林姬鼠的陰道能分泌一種黏液,只有超強運動能力的精子才能穿透它。針對這一防禦措施,機智而富有團隊精神的雄性小林姬鼠精子擺出「長蛇陣」,前仆後繼地對黏液層發起高破壞力的群攻,無私地幫助它們中的一個幸運兒最終完成受精任務。
最後就是最惡劣的「奧步」,刺穿式的交配方式。透過直接地把精子混入雌性體液,雄性臭蟲繞開了雌性精心設計的卵子防禦佈局。就算雌性在這個過程中受到了傷害,產生的後代數目因此變少,但從研究的統計數據來看,最終雄性依然在這個過程中扮演受益者的角色─健康的雌性固然好,但是如果不能產生「我」的後代,那就是一無所有。透過穿透式交配,雄性既阻止了雌蟲對他精子的抗拒,也扼殺了雌性天生挑三揀四的超強能力。

在雄性的角度上,這當然是一個成功的策略,以至於這種策略在動物界中不停地被重複演化出來。雄性吸口蟲(Myzostoma)的生殖器能分泌一種腐蝕性的酵素,從雌蟲的身上溶解出一個洞好讓他的精子得以進入;雄性巨烏賊能把其精液打包注入雌性的觸鬚中(儘管他們有時候會把「精液包」歪打正著地射到自己的腳裡XD);雄性捕潮蟲蛛則會撕咬雌性蜘蛛,然後通過針狀的生殖器刺札雌性蜘蛛,將精子從傷口注入雌蛛體內。
像寄椿這類昆蟲的雄性會胡亂地對雌性進行穿刺。在某些種類的動物中,不少雌性更上一層樓演化出減輕這樣傷害的構造,例如Ribaga所發現雌臭蟲腹部的「受精儲精器官」,透過提供方便的孔道導引雄蟲進入,以避免雄蟲不分青紅皂白的亂督。一些物種甚至棄置了本來的生殖系統,演化出一整套「副生殖系統」來引導精子進入卵巢,例如皮盲椿的雌蟲。
雄性臭蟲經常會不顧一切地撲向並刺扎一切的擋路者,甚至是其他物種的雌蟲,這常導致很多無辜血案發生,也在演化上促使這些倒楣的物種演化為其他的表型,避免被流彈強暴。雄性非洲臭蟲(Afrocimex constrictus)乾脆以暴制暴,雄蟲之間會用陰莖互戳,除了造成對方受傷外,也還是會把精子射進去落敗雄蟲體內,徹底地鄙視他。所以某些非洲臭蟲雄蟲又動了歪腦筋,不如讓演化把自己也加上了如雌蟲一般的「受精儲精器官」(沒錯,這就是偽娘)。
在這場生存博弈中,是沒有永遠的勝利者的─雄性可能會暫時占據上風,但是可想而知地雌性會馬上吹起反擊的號角。令人驚訝的是,一些非洲臭蟲的雌蟲逐步改變了其受精儲精器官的構造,改造地像那些偽娘身上的假「受精儲精器官」的樣式,以減少其他雄性的性騷擾事件。另外一些雌蟲則演化出消化精子的手段,並且把消化過後的能量拿來修補被穿刺的傷口,盡可能減輕傷害。
如你所見,真愛的道路上總是一波三折。在這場兩性生殖鬥爭中,雄性和雌性都在不停地繞著圈子打轉,永遠無法分出勝負,這或許是大自然開的一個惡毒的玩笑。不過想一想,其實這跟大家熟悉的蝙蝠跟蛾來來回回的攻防戰是類似的,都是屬於共演化的範疇,我們學演化的人稱作性別擷抗式共演化(sexual antagonistic coevolution)。
故事到這邊已經結束了,不過讓我來想想,或許,我們應該慶幸生為人類,才得以與這些可怕的戰場性事絕緣──多一點浪漫,才是擊破這個魔咒的唯一手段。

PS. 最後我們一起來大聲地對無脊椎動物們說:「你們根本不懂甚麼叫做愛!!!」
參考資料:
- 欢场即战场:那些有性无爱的动物们|果殼網 [2014-03-19]
- Rolanda Lange et al. (2013) Functions, diversity, and evolution of traumatic mating. Biological Review. (88)3: 585-601.
- Nikolai J. Tatarnic et al. (2014) Traumatic Insemination in Terrestrial Arthropods. Annu. Rev. Entomol. (59): 245-261.
- 果殼網原始編譯自The Conversation.的文章《Invertebrates inject a bit of romance during sex – by stabbing each other》
- 部分有肺類蝸牛的愛之飛鏢,射手比較有可能當爸爸。Ronald Chase & Katrina C. Blanchard. (2006) The snail’s love-dart delivers mucus to increase paternity. Proc. R. Soc. B. (273): 1471–1475
- 扁蟲的揮舞老二之戰。N. K. Michiels & L. J. Newman. (1998) Sex and violence in hermaphrodites. Nature. (391): 647
- 北美此種豆娘(Ebony Jewelwing)的雄性生殖器舀精功能。Jonathan K. Waage. (1979) Dual Function of the Damselfly Penis: Sperm Removal and Transfer. Science. 203(4383): 916-918.
- 雌性野禽會排除弱勢雄性的精液。T. Pizzari & T. R. Birkhead. (2000) Female feral fowl eject sperm of subdominant males. Nature. (405): 787-789.
- 雄性吸口蟲(Myzostoma)的生殖器分泌物溶掉雌蟲腹部。I. Eeckhaut & M. Jangoux. (1991) Fine structure of the spermatophore and intradermic penetration of sperm cells inMyzostoma cirriferum (Annelida, Myzostomida). Zoomorphology. (111)1: 49-58.
- 雄性小林姬鼠精子合作形成「長蛇陣」。Harry Moore et al. (2002) Exceptional sperm cooperation in the wood mouse. Nature. (418): 174-177
- 雄果蠅在精子中加料,使雌果蠅壽命減短。Brett Holland & William R. Rice. (1999) Experimental removal of sexual selection reverses intersexual antagonistic coevolution and removes a reproductive load. PNAS. (96) 9: 5083-5088
- 雌性臭蟲消化不要的精子來保養身體。Klaus Reinhardt et al. (2009) Ejaculate components delay reproductive senescence while elevating female reproductive rate in an insect. PNAS. (106)51: 21743-21747







































