露兜樹象鼻蟲的身世之考察——分類學家偵探事件簿(四)
一般大眾或甚至其他領域生物學家們,對於基礎生物分類學家的刻板印象,無非是常常在顯微鏡下進行形態解剖比較來鑑定物種、描述並發表新物種,或者常常東跑西跑去採集標本,頂多是抽取遺傳物質進行 DNA 分析。然而一位稱職的分類學家,為了搞清楚物種學名的分類地位,將整個命名系統修訂成一個穩定並適合大家使用的狀態,往往需要做大量的歷史文獻,造訪各大博物館並進行模式標本考察,其中的繁瑣和複雜程度,往往令人出乎意料。
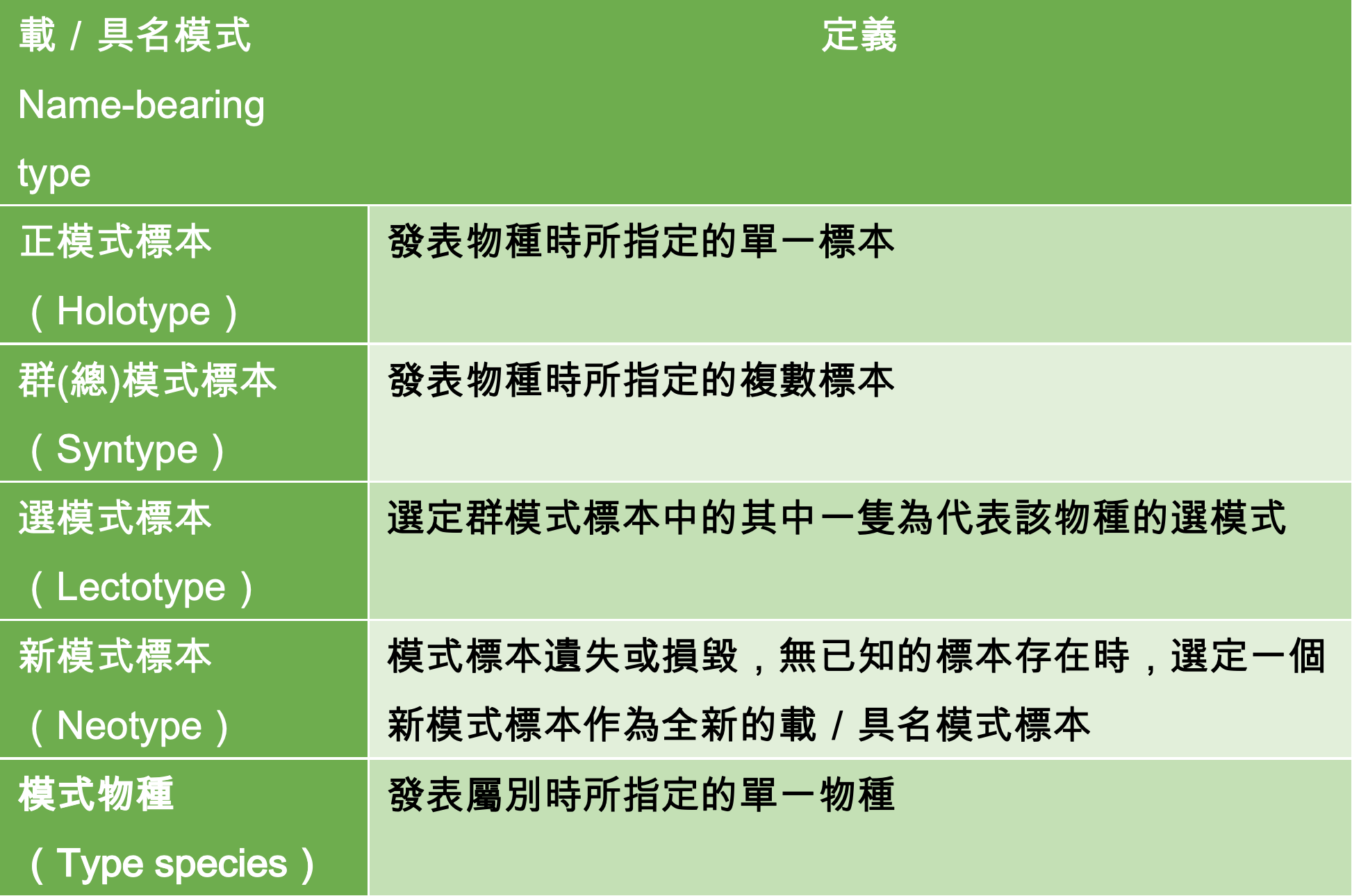
再讓我們複習一次模式標本是什麼和其重要性?
如果有閱覽過這系列的文章便會很清楚的知道,模式標本是物種發表時的實體存證,是學者對分類地位有疑慮時,用以判別的客觀證據。每個物種都有其模式標本,而每個屬也有其模式物種,是判定該屬別的決定性物種,模式種和模式標本是進行物種與屬別層級的基礎分類研究時,不可或缺的重要資訊。
這個故事的主角是一類來自南亞和東南亞的露兜樹象鼻蟲,本文將講述其模式標本和背後歷史脈絡的考察,以及我們對於分類處理過程的案例分享。
分布於南亞、東南亞的露兜樹象鼻蟲和研究緣起
露兜樹科(Pandanus )為分布於東半球的亞熱帶及熱帶地區的灌木或喬木植物,其中林投(Pandanus tectorius )具有抗風、耐鹽的特性,是常見的海岸防風定砂植物,而俗稱斑蘭葉(pandan)的七葉蘭(Pandanus amaryllifolius ),則是東南亞常見的料理與糕點製作材料,而南亞和東南亞的露兜樹上棲息著一群黑色扁平的小型象鼻蟲——露兜樹象鼻蟲(Lyterius )。
露兜樹是東半球的亞熱帶及熱帶地區的灌木或喬木植物。(攝/B.navez from Wikipedia)
小小扁扁的露兜樹象鼻蟲(Lyterius )是與露兜樹有伴生關係的特別物種。(圖/論文原文)
而故事的緣起可追溯到 2022 年,當時筆者正在澳洲進行博士論文題目「澳洲蘇鐵授粉象鼻蟲的多樣性與演化」的研究,我們意外地發現澳洲的蘇鐵授粉象鼻蟲與東南亞產的露兜樹象鼻蟲親緣關係接近,因此我們便想進一步探究本類群的分類。在我們初步搜索模式標本時,我們驚奇地發現德國象鼻蟲學者延斯・普雷納博士 Dr. Jens Prena 似乎曾經有研究過這類象鼻蟲,出於好奇,我們聯繫了普雷納博士,進而開啟了本類群錯綜複雜的分類歷史考察之旅。
露兜樹象鼻蟲分類研究的現存問題
首先露兜樹象鼻蟲的分類問題分成兩個面向,一個是屬別層級的,而另一個是物種層級的。屬別層級的問題比較簡單,我們發現露兜樹象鼻蟲屬有三個相關的屬別,分別為 Lyterius Schönherr, 1844、Barisoma Motschulsky, 1863 和 Plaxes Pascoe, 1885,根據牠們形態的相似性和地理分布的重疊,我們認為牠們應該被合併成單一屬別,也就是說只要我們確認三個屬別的模式種都是屬於同一個屬別後,那自然我們就能依照優先權原則,把 1863 年發表的 Barisoma 和 1885 年發表的 Plaxes 處理為最早發表的 Lyterius 的同物異名。
但是!分類學研究最困難的就是這個但是!
我們雖然追蹤到 Barisoma 和 Plaxes 的模式種和其模式標本,但是 Lyterius 的模式種問題,卻將這個研究的難度拉向了另一個層面——也就是物種層級的問題。
模式標本來源和流向超級複雜的 Lyterius
Lyterius 這個屬別是由瑞典昆蟲學家卡爾・約翰・舍恩赫爾(Carl Johan Schönherr)於 1844 年所提出,並以 Rhynchaenus musculus Fabricius, 1802,這個 1802 年由丹麥昆蟲學家約翰・克里斯蒂安・法布里丘斯(Johan Christian Fabricius )所發表的種類作為模式物種。他的合作對象瑞典昆蟲學家卡爾・亨利克・博赫曼(Carl Henrik Boheman)也在同一本書中使用了 Lyterius musculus (Fabricius, 1802) 這個學名組合,同時他將德國昆蟲學家弗里德里希・韋伯(Friedrich Weber)在 1802 年所描述的 Curculio abdominalis Weber, 1801 也拉進這個屬別,學名組合變成 Lyterius abdominalis (Weber, 1801) ,並且描述一個菲律賓的新物種 Lyterius instabilis Boheman in Schönherr, 1844 。這其中最為複雜難解的,便是 Lyterius musculus (Fabricius, 1802) 和 Lyterius abdominalis (Weber, 1801) 之間的關係了,因為這兩個物種的模式標本來源,都源自於達戈貝爾特・達爾多夫 Dagobert Karl von Daldorff 這位在俄羅斯出生,擁有德裔血統的丹麥博物學家,在 18 世紀末葉任職丹麥東印度公司時,於 1795 年在蘇門答臘的一次採集。
除了我們常常聽到的荷蘭、英國東印度公司,丹麥也曾經創立了東印度公司。(攝/Wikipedia)
根據我們對於 19 世紀初期的歐洲甲蟲分類歷史文獻的爬梳,達爾多夫在蘇門答臘的標本被帶回歐洲後,應該至少被他贈與或交換給五位學者或機構,而這五位學者就包含剛剛提到的德國昆蟲學家弗里德里希・韋伯(Friedrich Weber),以及丹麥昆蟲學家約翰・克里斯蒂安・法布里丘斯(Johan Christian Fabricius),這兩位顯然同時對這批標本進行分類學研究。
令人存疑的 Lyterius abdominalis 和 Lyterius musculus
因此第一個疑點就是,韋伯和法布里丘斯分別在 1801 年和 1802 年用達爾多夫所採集的同一批蘇門答臘象鼻蟲標本,發表了後來在 1844 年被博赫曼放在同一個屬別的物種 Lyterius abdominalis 和 Lyterius musculus ,這讓人很難不懷疑,這兩個名字會不會根本就是同一個物種,這在當年資訊不流通、分類研究還很粗淺的年代,是非常容易發生的事情。
而支持這樣想法的關鍵則有二,首先德國昆蟲學家約翰・卡爾・威廉・伊利格(Johann Karl Wilhelm Illiger)其實在 1805 年的著作中,就已經提出這兩個物種是同一個物種的論點了,然而這項分類處理卻被博赫曼在 1844 年的著作中,不明地忽略了。雖然博赫曼不小心遺漏了伊利格的分類處理,他卻也在看過兩種的模式標本後,在他那 1844 年的著作中,提出了兩個物種只不過是同一個物種的雄蟲和雌蟲的猜想,然而因為他手邊就只有兩隻標本,一隻是雄的 Lyterius abdominalis ,一隻是雌的 Lyterius musculus ,因此他無法下這個決定情有可原,而我們如今已經知道露兜樹象鼻蟲有很明顯的雌雄二形性,雄蟲的口喙比較短,且足部的前腳腿節有明顯的突起,博赫曼的猜想不證自明。
總而言之,從上述的歷史文獻爬梳,我們可以從
韋伯和法布里丘斯研究的都是同一批蘇門答臘採集的標本
同時代的伊利格和後來的博赫曼都直接或間接的認為 Lyterius abdominalis 和 Lyterius musculus 是同一個物種
來推斷,這兩個種類很有可能是同一個種類!
瑞典昆蟲學家卡爾・亨利克・博赫曼。(攝/Wikipedia)
找不到模式標本啊!
在爬梳大量文獻後,我們同時也造訪歐陸各大標本蒐藏去尋找這些物種的模式標本下落。我們很幸運的在德國基爾的動物學博物館找到兩隻 Lyterius musculus 的總/群模式標本。然而,在尋找 Lyterius abdominalis 模式標本的過程中卻碰了壁,不管是文獻還是實際探訪,幾乎都找不到韋伯收藏的下落,韋伯所發表的模式標本有極大的可能已經遺失了,那要怎麼辦呢?
分類學家的決策
雖然沒辦法找到 Lyterius abdominalis 的模式標本,然而我們從以上的間接證據,可以合理相信 Lyterius abdominalis 和 Lyterius musculus 就是同一個物種。為了最適當的處理分類議題,穩定整個分類命名系統。我們使用了一個技術性的分類學處理,首先我們指定了 Lyterius musculus 的選模式標本,並且我們將「這一個」標本,再次的指定為 Lyterius abdominalis 的新模式標本,這個時候,這兩個學名便產生了動物命名法規上所謂的「客觀同物異名(objective synonym)」關係,相較於分類學家自行主觀認定的同物異名(主觀同物異名 subjective synonym ),客觀同物異名指的是用同一個標本發表不同學名的狀況,這樣這兩個名字無庸置疑的是同物異名關係,只有最早被發表的名字有優先權,因此我們的 Lyterius abdominalis (Weber, 1801) 獲得了優先被使用的地位,也成為露兜樹象鼻蟲屬的模式種。經由這一波操作,我們確立了 Lyterius 的模式和包含的物種,也因此我們終於能進一步處理剛剛提到的 Barisoma 和 Plaxes 的同物異名,最後我們可以大聲的說:露兜樹象鼻蟲屬的學名是 Lyterius Schönherr, 1844 !
番外篇的 Plaxes 模式標本調查
另外一方面,我們在調查 Plaxes 的模式標本時,也發現到其模式種 Plaxes impar Pascoe, 1885 的總/群模式標本散落在英國倫敦自然史博物館、德國柏林自然史博物館、德國德勒斯登森肯堡博物館、義大利熱拿亞自然史博物館、澳洲國立昆蟲館,幾乎涵蓋了半個地球。這些標本可以分為來自婆羅洲砂拉越和蘇門答臘的標本,採自砂拉越的標本無疑是一個獨立的物種,我們也指定砂拉越的總/群模式標本為本種選模式標本。而來自蘇門答臘的標本,無獨有偶地都和 Lyterius abdominalis 是同一個物種,顯然這個物種在蘇門答臘當地是個常見的物種,這又再次加強我們上面提到的,達爾多夫所採集的同一批蘇門答臘象鼻蟲標本應該就只有一種露兜樹象鼻蟲的推測。
這個研究重新梳理了露兜樹象鼻蟲的分類歷史並考察了歷史文獻和模式標本,最終作出了適宜的分類學處理,為亞洲地區的象鼻蟲研究推進了一步。
本論文日前已經線上刊載於《動物分類群 Zootaxa 》
此文響應 PanSci 「自己的研究自己分享」,以增進眾人對基礎科學研究的了解。
參考資料
Prena, J., Hsiao, Y., Oberprieler, R.G. (2023) New combinations and synonymies in the weevil genus Lyterius Schönherr (Coleoptera, Curculionidae), with a conspectus of historical works on Daldorff’s Sumatran beetles. Zootaxa 5380(1): 26-36. https://doi.org/10.11646/zootaxa.5380.1.2