長期以來,動物著色策略一直都是博物學家和哲學家感興趣的話題,甚至在 2000 多年前亞里士多德的《動物史》中也早已出現過相似的議題[34]。在過去的幾個世紀中,科學家們開始認真探索動物顏色是如何產生的[45],並闡明它們可能的功能[11, 12, 57]。
其中哺乳類的顏色一般都被認為是比較單調的[8],印象中雖然有看過五色鳥、變色龍和箭毒蛙等顏色鮮豔的動物了,但相比之下五顏六色的哺乳類確實難以想像,這不禁讓人思考是不是有哪些演化環節造成如此的差異?
本篇文章將以多元的觀點作切入,一步步探討為什麼哺乳類著色(coloration)會不夠豐富,並且梳理其中相關議題的脈絡。
什麼影響著色策略?
首先釐清這個問題必需要分成幾個層次。
- 發色機制
除了生物螢光(biofluorescence)外,所有動物顏色都是由兩種主要機制所產生的:(1) 色素著色以及 (2) 結構著色[16]。
舉例來說,藍色顏料在自然界其實是稀缺的,大部分鳥類的藍色羽毛(如台灣藍鵲)就是利用結構著色,而不是藍色色素[2, 35, 42]。
其中生物色素如類胡蘿蔔素(carotenoid),吸收光譜約為 400–500nm,能在動物體內產生紅色至黃色,而黑色素在所有可見波長範圍內都表現出高吸光度,真黑素(eumelanin)產生黑色至金色;棕黑素(phaeomelanin)產生淡黃色至淡紅色[21, 28, 36]。
而不同於其他脊椎動物,目前除了洪都拉斯白蝙蝠(Ectophylla alba)能夠在皮膚中累積類胡蘿蔔素以外[22],大部分哺乳類都只能在皮膚與毛髮中產生黑色素[7]。
- 視覺系統
再來必須意識到的是,我們人類所感知到的顏色與其他動物自己所感知到的顏色是兩回事。
生物的體色對於各式各樣的行為和演化研究非常重要,這些信號包括隱蔽(crypsis)、擬態(mimicry)、警戒色(warning coloration)、花果著色以及雌雄二型性(sexual dimorphism)等[4, 18]。
對於這些特徵來說,真正重要的是這些信號的目標受眾的色覺(color vision),無論是捕食者、傳粉者還是潛在配偶等[23]。所以有人就認為顏色不是物體的固有屬性,而是能感知它的生物體視覺系統屬性[17]。
換句話說,根據動物光感受器的吸收光譜的不同,從物體反射的波長光譜將被感知為不同的顏色[17, 24]。而關於色覺的差異,不同種類的視蛋白(opsins)就代表著不同的吸收光譜,然而在當代主要脊椎動物群中,四種視蛋白(代表四色視覺)是廣泛存在於各種鳥類、魚類和爬蟲類中的,但意外的是大部分哺乳類卻是雙色視覺[6]。
重要的演化事件——夜行瓶頸(nocturnal bottleneck)
所以是什麼造成哺乳類與鳥類、爬蟲類的視覺差異?
- 時間背景
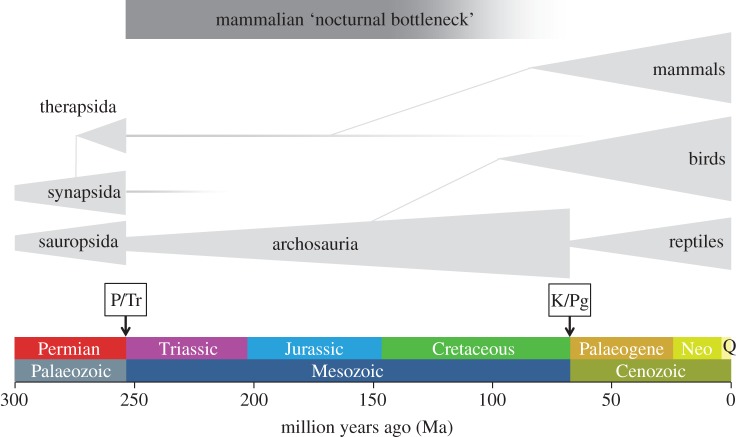
時間拉回到 2.5 億年前左右,哺乳動物最早是從三疊紀的獸孔目祖先演化而來,根據對化石證據的分析推斷,這些幾乎是夜行性的小型食蟲動物[31],關於這個現象有些議題會專注於討論視覺[50, 58],而有些會談地更全面[48, 60]。具體的夜行瓶頸假說最初是由Menaker等人制定[20, 43],內容主要是推測一些類哺乳爬蟲類動物(合弓綱)在 P/Tr 事件(二疊紀/三疊紀滅絕事件)中倖存下來,並大約發展了 1.45 億年後形成了所謂的「中生代哺乳類」(圖1)。
- 生態背景
夜行瓶頸假說認為早期哺乳動物在中生代期間會面臨到日行性爬蟲類(主龍類,如恐龍)的競爭[10, 29, 43, 58],而這一系列的適應性變化使早期哺乳動物能不用受太陽輻射和環境溫度的限制下在夜間進行活動,並促進其內溫系統的發展[23]。相對地,長期適應夜間活動可能使光感受器(photoreceptor)發生巨大變化,包括未充分利用之光感受器功能與紫外線保護機制的喪失[23]。

- 視覺光感受系統
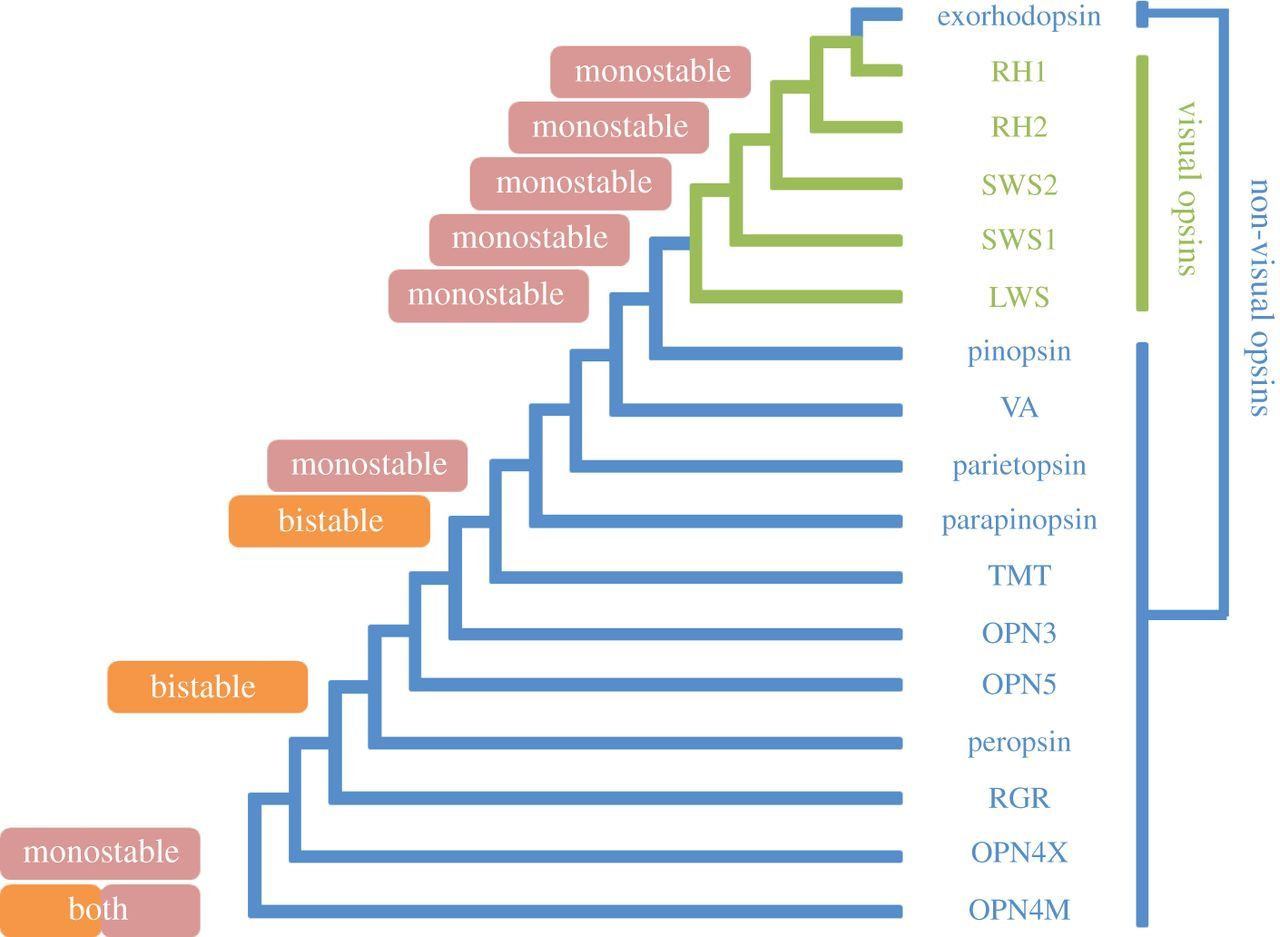
視錐細胞(cone cells)和視桿細胞(rod cells)內的感光色素皆是由視蛋白組成,而視蛋白基因可分為五個亞型:(sws1、sws2、rh1、rh2 和 lwS)[13, 59],它們的接收光譜範圍可以參考(圖2)。
儘管可能所有五種視蛋白基因早期都存在於合弓綱中,但隨著真獸下綱(eutherian)的演化輻射,視蛋白基因的多樣性就開始急劇減少了[13](圖1)。首先所有哺乳類祖先都丟失了 rh2 基因,雖然單孔目動物(鴨嘴獸和針鼴等)還是有保留著 sws1 基因,但是卻失去其功能,隨即真獸下綱也丟失了 sws2 基因,只剩 sws1 和 lwS 能表達[13],導致至今大部分的哺乳類都屬於雙色視覺 [14, 56](表1)。
- 非視覺光感受系統
儘管在整個哺乳動物演化過程中,色覺的變化是被認為神經生態適應最經典的例子之一,但事實上夜行瓶頸在塑造非視覺光感受(non-visual photoreception)方面,也發揮了相當的影響力[23]。其中黑視蛋白 (melanopsin,Opn4) 就扮演了非常重要的角色,它負責晝夜節律的光誘導及調節其他生理反應[23],例如:身體色素沉著的變化[51]、瞳孔收縮[38]、褪黑激素抑制[39]、睡眠誘導[40]及暗光運動[19](dark photokinesis[註2]) 等。
黑視蛋白有 Opn4X 和 Opn4M 這兩種基因(Opn4M 是 Opn4X 的基因重複變體),它們廣泛存在於非哺乳動物脊椎動物中,但 Opn4X 在早期哺乳動物演化中丟失[3, 15](表1)。至於這兩個基因的差異並不明顯,主要在於組職表達模式的不同[26],且至今仍然沒有完全理解為什麼所有現存哺乳動物中保留的是 Opn4M 而非 Opn4X[23]。

其他哺乳類的著色特例
- 舊世界猴
約 8–9 千萬年前第一批靈長類動物出現了[5, 54],雖然這些早期靈長類通常也被認為是夜行性的[41],但隨後有許多靈長類動物譜系開始漸漸適應白天活動。直至大約 3–4 千萬年前,不同於其他哺乳類,所有狹鼻小目(catarrhine primates,包含舊世界猴、猿類與人類)都藉由基因重複多產生了一個 LWS 基因[44, 9],而這種革新開始提供靈長類重新擁有了三色視覺的能力。例如山魈因此得以演化出藍色的臉譜,但牠們利用的原理是相干散射(coherently scattering)[49],而不是單純的色素。
2022/07/06 編按:
多出來的 LWS 基因只是感測綠色,和藍色的山魈臉譜沒關係,前後並沒有因果關係。

- 能生物螢光的哺乳類
生物螢光簡單來說就是生物將短波長的光吸收後並重新發射成長波長的光,與舊印象不同,有趨勢顯示越來越多哺乳類物種被發現能夠進行生物螢光,雖然生物螢光的生態功能與起源目前還有爭議,但可能與中新世古氣候及物種的生活型態具有密切關係[25]。
目前已知的在紫外線下具有生物螢光毛皮的哺乳類物種中,全部都是夜行性的,其中包括:鴨嘴獸[1]、美洲飛鼠屬 ( Glaucomys spp.)[32]、跳兔(Pedetes capensis)[46]和一些負鼠科 ( Didelphidae)物種[47]。
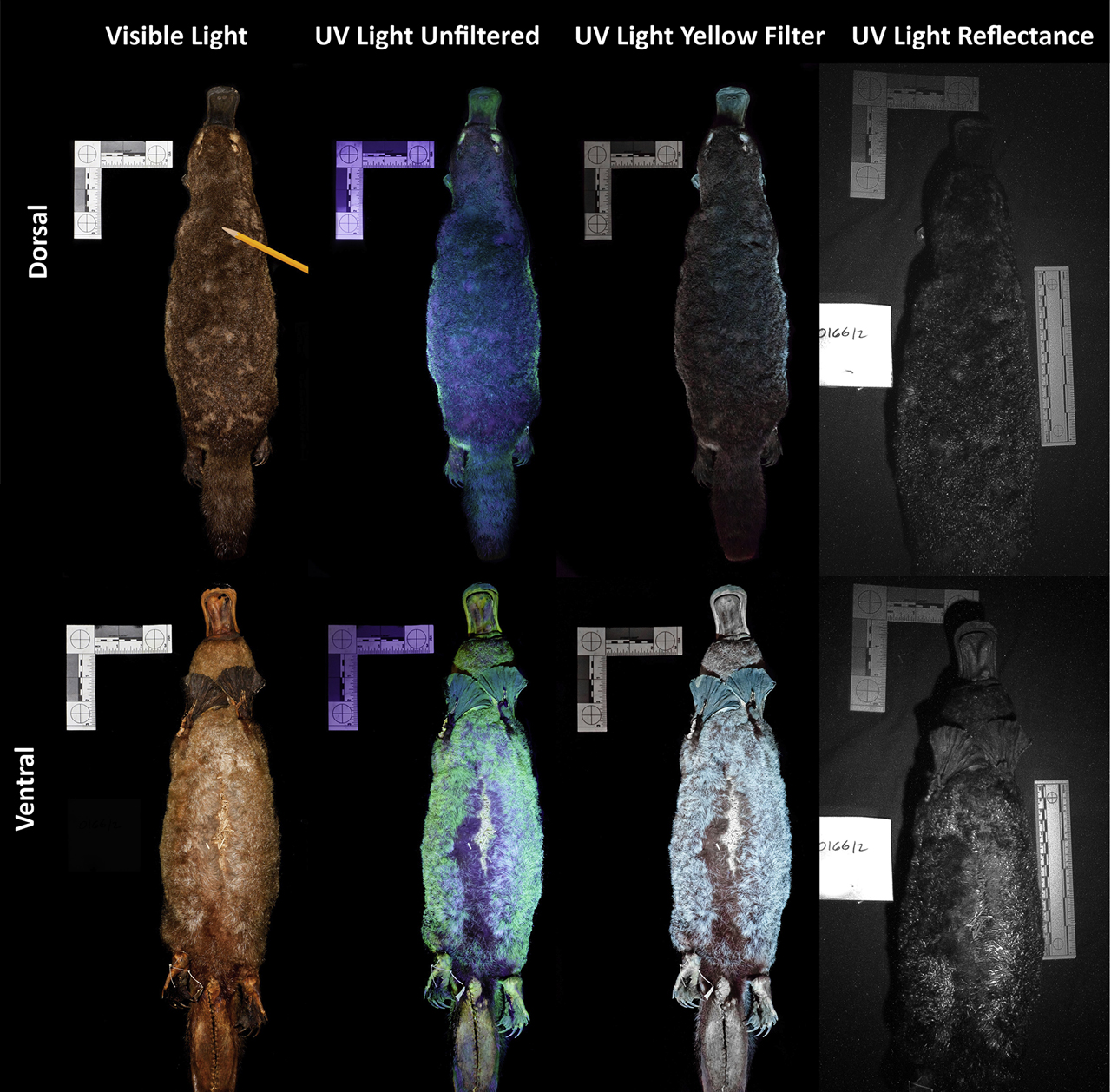
- 非洲金鼴
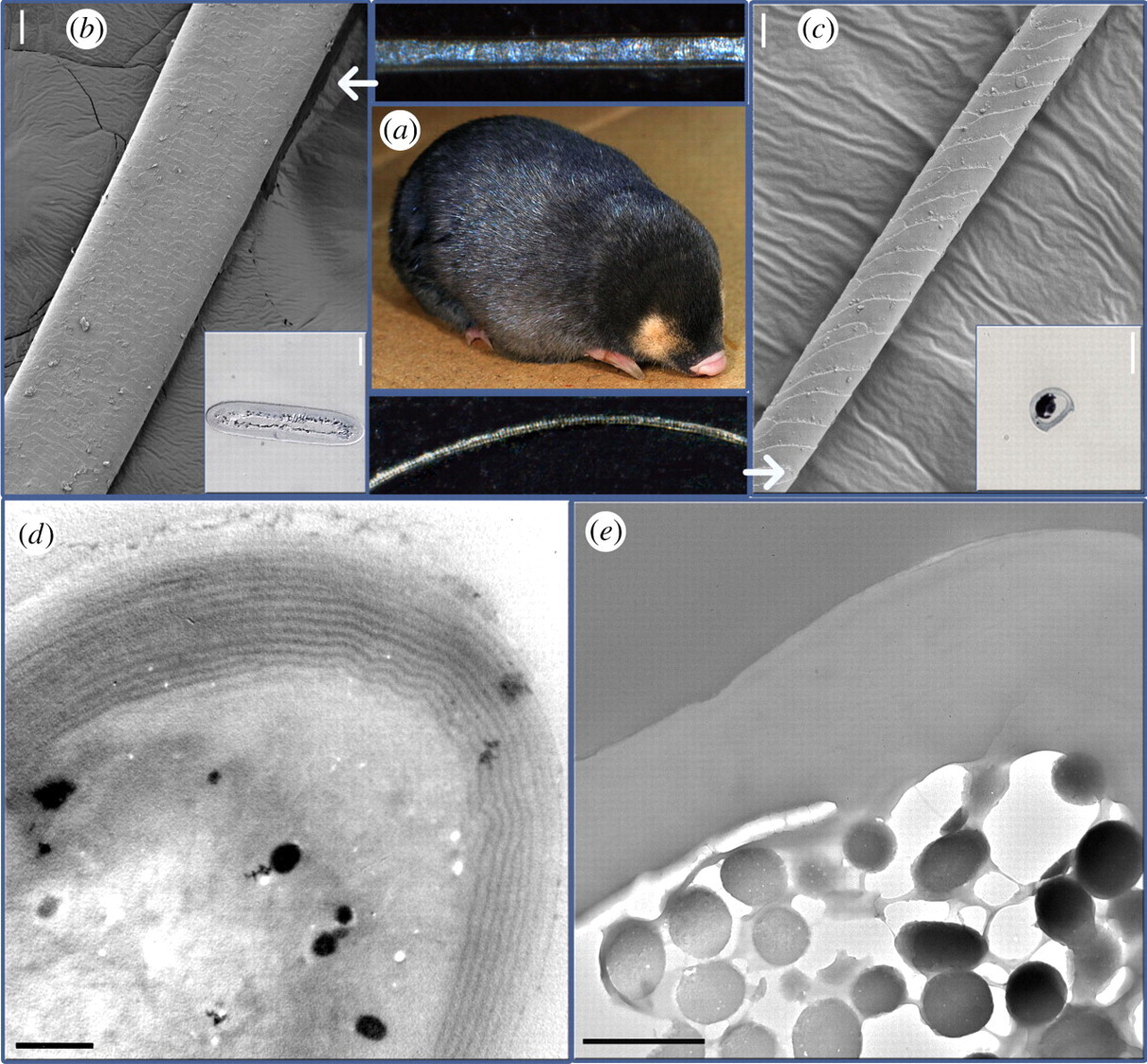
非洲金鼴(Amblysomus hottentotus、Amblysomus septentrionals、Chrysochloris asiatica & Eremitalpa granti)是一群撒哈拉以南特有的視力退化食蟲性穴居動物[33],牠們的毛髮具有特殊的虹彩(iridescent),能反射出藍綠色的高光,並且在電子顯微鏡下Snyder等人發現這些槳狀毛髮的超微結構[53],這構造除了能成色外還具有增加毛鱗耐磨性與減少摩擦力等功能,他們假設:因為缺乏視覺能力,非洲金鼴的虹彩既不是性擇的產物,也不可能起到偽裝的作用,唯一的可能就是毛髮在這種結構下能夠幫助非洲金鼴在地下移動時,減少在泥沙中的湍流,而毛髮的虹彩可能就只是演化下的副產品。
哺乳類的著色並沒有想像中的單調
雖然本篇的先入為主認為哺乳類的著色就是不豐富的,但如果論及花紋以及圖案了話,也有觀點是認為哺乳類的著色其實是很多樣的[8],況且如上面所提及的案例,就算以我們人類的視角,也有越來越多證據顯示某些哺乳類的顏色也沒那麼單調。正如前面所說的對於動物的著色而言,真正重要的是訊息的接受者,所以如果侷限於人類的視角了話,其實是難以了解其全貌的。
最後要提醒的是,即使夜行瓶頸影響了之後哺乳類的視覺系統,很多證據也顯示視覺與著色之間是有共演化關係的,特別是那些具有婚姻色或雌雄二型性的物種,但是對於隱蔽與偽裝色等來說,證據其實是有限的[37,55],更何況有如同非洲金鼴一樣的個案存在[53]。所以總體而言,這算是一個還滿複雜的議題,某些時候在宏觀演化學上不一定有所謂的必然[27],演化並不是說一定會這樣發展或那樣發展,更多的其實是要看這些生物在一定時空間尺度下要如何應對及適應。
註解
參考資料
- Anich, P. S., Anthony, S., Carlson, M., Gunnelson, A., Kohler, A. M., Martin, J. G., & Olson, E. R. (2020). Biofluorescence in the platypus (Ornithorhynchus anatinus). Mammalia, 85(2), 179–181. doi: 10.1515/mammalia-2020-0027
- Bagnara, J. T., Fernandez, P. J., & Fujii, R. (2007). On the blue coloration of vertebrates. Pigment Cell Research, 20(1), 14–26. doi:10.1111/j.1600-0749.2006.00360.x
- Bellingham, J., Chaurasia, S. S., Melyan, Z., Liu, C., Cameron, M. A., Tarttelin, E. E., Iuvone, P. M., Hankins, M. W., Tosini, G., & Lucas, R. J. (2006). Evolution of Melanopsin Photoreceptors: Discovery and Characterization of a New Melanopsin in Nonmammalian Vertebrates. PLoS Biology, 4(8), e254. doi: 10.1371/journal.pbio.0040254
- Bennett, A. T. D., Cuthill, I. C., & Norris, K. J. (1994). Sexual Selection and the Mismeasure of Color. The American Naturalist, 144(5), 848–860. doi: 10.1086/285711.
- Bininda-Emonds, O. R. P., Cardillo, M., Jones, K. E., MacPhee, R. D. E., Beck, R. M. D., Grenyer, R., Price, S. A., Vos, R. A., Gittleman, J. L., & Purvis, A. (2007). The delayed rise of present-day mammals. Nature, 446(7135), 507–512. doi: 10.1038/nature05634
- Bowmaker, J. K. (2008). Evolution of vertebrate visual pigments. Vision Research, 48(20), 2022–2041. doi: 10.1016/j.visres.2008.03.025
- Bradley, B. J., & Mundy, N. I. (2008). The primate palette: The evolution of primate coloration. Evolutionary Anthropology: Issues, News, and Reviews, 17(2), 97–111. doi: 10.1002/evan.20164
- Caro, T. (2005). The adaptive significance of coloration in mammals. BioScience 55(2), 125–136. doi: 10.1641/0006-3568(2005)055[0125:TASOCI]2.0.CO;2
- Carvalho, L. S., Pessoa, D. M. A., Mountford, J. K., Davies, W. I. L., & Hunt, D. M. (2017). The Genetic and Evolutionary Drives behind Primate Color Vision. Frontiers in Ecology and Evolution, 5(34). doi: 10.3389/fevo.2017.00034
- Crompton, A. W. (1980). Biology of the earliest mammals. In K. Schmidt-Nielsen, L. Bolis, & C. R. Taylor (Eds.), Comparative Physiology: Primitive Mammals (pp. 1–12). New York, NY: Cambridge University Press.
- Darwin, C. (1859). On the origin of species by means of natural selection. London, UK: Murray.
- Darwin, C. (1871). The descent of man and selection in relation to sex. London, UK: Murray.
- Davies, W. I, Collin, S. P., & Hunt, D. M. (2012). Molecular ecology and adaptation of visual photopigments in craniates. Molecular Ecology, 21(13), 3121–3158. doi: 10.1111/j.1365-294X.2012.05617.x
- Davies, W. L., Carvalho, L. S., Cowing, J. A., Beazley, L. D., Hunt, D. M., & Arrese, C. A. (2007). Visual pigments of the platypus: A novel route to mammalian colour vision. Current Biology, 17(5), R161–R163. doi: 10.1016/j.cub.2007.01.037
- Davies, W. L., Hankins, M. W., & Foster, R. G. (2010). Vertebrate ancient opsin and melanopsin: divergent irradiance detectors. Photochemical & Photobiological Sciences, 9(11), 1444. doi: 10.1039/c0pp00203h
- Doucet, S. M., & Meadows, M. G. (2009). Iridescence: a functional perspective. Journal of The Royal Society Interface, 6(Suppl_2), S115–S132. doi: 10.1098/rsif.2008.0395.focus
- Endler, J. A. (1978). A Predator’s View of Animal Color Patterns. Journal of Evolutionary Biology, 11(3), 319–364. doi: 10.1007/978-1-4615-6956-5_5
- Endler, J. A. (1990). On the measurement and classification of colour in studies of animal colour patterns. Biological Journal of the Linnean Society, 41(4), 315–352. doi: 10.1111/j.1095-8312.1990.tb00839.x.
- Fernandes, A., Fero, K., Arrenberg, A., Bergeron, S., Driever, W., & Burgess, H. (2012). Deep Brain Photoreceptors Control Light-Seeking Behavior in Zebrafish Larvae. Current Biology, 22(21), 2042–2047. doi: 10.1016/j.cub.2012.08.016
- Foster, R. G., & Menaker, M. (1993). Circadian Photoreception in Mammals and Other Vertebrates. Light and Biological Rhythms in Man, 73–91. doi: 10.1016/b978-0-08-042279-4.50009-1
- Fox, D. L. (1953). Animal Biochromes and Structural Colours. Amsterdam University Press.
- Galván, I., Garrido-Fernández, J., Ríos, J., Pérez-Gálvez, A., Rodríguez-Herrera, B., & Negro, J. J. (2016). Tropical bat as mammalian model for skin carotenoid metabolism. Proceedings of the National Academy of Sciences, 113(39), 10932–10937. doi: 10.1073/pnas.1609724113
- Gerkema, M. P., Davies, W. I. L., Foster, R. G., Menaker, M., & Hut, R. A. (2013). The nocturnal bottleneck and the evolution of activity patterns in mammals. Proceedings of the Royal Society B: Biological Sciences, 280(1765), 20130508–20130508. doi: 10.1098/rspb.2013.0508
- Gerl, E. J., & Morris, M. R. (2008). The Causes and Consequences of Color Vision. Evolution: Education and Outreach, 1(4), 476–486. doi: 10.1007/s12052-008-0088-x
- Gray, R., & Karlsson, C. (2022, February 6). 101 years of biofluorescent animal studies: trends in literature, novel hypotheses, and best practices moving forward. doi: 10.32942/osf.io/ub6yn.
- Hauzman, E., Kalava, V., Bonci, D., & Ventura, D. F. (2019). Characterization of the melanopsin gene (Opn4x) of diurnal and nocturnal snakes. BMC evolutionary biology, 19(1), 174. doi: 10.1186/s12862-019-1500-6
- Haufe, C. (2015). Gould’s Laws. Philosophy of Science, 82(1), 1–20. doi: 10.1086/678979
- Hill, G. E. (2010). National Geographic Bird Coloration (Illustrated ed.). National Geographic.
- Hut, R. A., Kronfeld-Schor, N., van der Vinne, V., & de la Iglesia, H. (2012). In search of a temporal niche. Progress in Brain Research, 199, 281–304. doi: 10.1016/B978-0-444-59427-3.00017-4
- Jacobs, G. H. (2009). Evolution of colour vision in mammals. Philosophical Transactions of the Royal Society B: Biological Sciences, 364(1531), 2957–2967. doi: 10.1098/rstb.2009.0039
- Kemp, T. S. (2005). The origin and evolution of mammals Oxford. UK: Oxford University Press.
- Kohler, A. M., Olson, E. R., Martin, J. G., & Anich, P. S. (2019). Ultraviolet fluorescence discovered in New World flying squirrels (Glaucomys). Journal of Mammalogy, 100(1), 21–30. doi: 10.1093/jmammal/gyy177
- Kuyper, M. A. (1985). The ecology of the golden mole Amblysomus hottentotus. Mammal Review, 15(1), 3–11. doi: 10.1111/j.1365-2907.1985.tb00379.x
- Lennox, J. G. (2002). Aristotle: On the Parts of Animals I-IV (Clarendon Aristotle Series) (1st ed.). Clarendon Press.
- Liao, S. F., Yao, C. Y., & Lee, C. C. (2015). Measuring and modeling the inconspicuous iridescence of Formosan blue magpie’s feather (Urocissacaerulea). Applied Optics, 54(16), 4979. doi: 10.1364/AO.54.004979
- Lichtenthaler, H. K., & Buschmann, C. (2001). Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Current Protocols in Food Analytical Chemistry, 1(1), F4.3.1–F4.3.8. doi: 10.1002/0471142913.faf0403s01
- Lind, O., Henze, M. J., Kelber, A., & Osorio, D. (2017). Coevolution of coloration and colour vision? Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1724), 20160338. doi: 10.1098/rstb.2016.0338
- Lucas, R. J., Douglas, R. H., & Foster, R. G. (2001). Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Nature Neuroscience, 4(6), 621–626. doi: 10.1038/88443
- Lucas, R. J., & Foster, R. G. (1999). Neither Functional Rod Photoreceptors nor Rod or Cone Outer Segments Are Required for the Photic Inhibition of Pineal Melatonin*. Endocrinology, 140(4), 1520–1524. doi: 10.1210/endo.140.4.6672
- Lupi, D., Oster, H., Thompson, S., & Foster, R. G. (2008). The acute light-induction of sleep is mediated by OPN4-based photoreception. Nature Neuroscience, 11(9), 1068–1073. doi: 10.1038/nn.2179
- Martin, R. D., & Ross, C. F. (2006). The Evolutionary and Ecological Context of Primate Vision. The Primate Visual System, 1–36. doi: 10.1002/0470868112.ch1
- Mason, C. W. (1923). Structural Colors in Feathers. I. The Journal of Physical Chemistry, 27(3), 201–251. doi: 10.1021/j150228a001
- Menaker, M., Moreira, L., & Tosini, G. (1997). Evolution of circadian organization in vertebrates. Brazilian Journal of Medical and Biological Research, 30(3), 305–313. doi: 10.1590/s0100-879×1997000300003
- Nathans, J., Thomas, D., & Hogness, D. S. (1986). Molecular Genetics of Human Color Vision: The Genes Encoding Blue, Green, and Red Pigments. Science, 232(4747), 193–202. doi: 10.1126/science.2937147
- Newton, I., Cohen, B. I., Einstein, A., & Whittaker, E. (2012). Opticks: Or a Treatise of the Reflections, Refractions, Inflections & Colours of Light-Based on the Fourth Edition London, 1730. Dover Publications.
- Olson, E. R., Carlson, M. R., Ramanujam, V. M. S., Sears, L., Anthony, S. E., Anich, P. S., … Martin, J. G. (2021). Vivid biofluorescence discovered in the nocturnal Springhare (Pedetidae). Scientific Reports, 11(1). doi: 10.1038/s41598-021-83588-0
- Pine, R. H., Rice, J. E., Bucher, J. E., Tank, D. H. J., & Greenhall, A. M. (1985). Labile pigments and fluorescent pelage in didelphid marsupials. Mammalia, 49(2). doi: 10.1515/mamm.1985.49.2.249
- Pough, H. F., Heiser, J. B., & McFarland, W. N. (1989). Vertebrate Life (3rd ed.). Macmillan Coll Div.
- Prum, R. O. (2004). Structural colouration of mammalian skin: convergent evolution of coherently scattering dermal collagen arrays. Journal of Experimental Biology, 207(12), 2157–2172. doi: 10.1242/jeb.00989
- Schwab, I. R. (2012). Evolutions witness: how eyes evolved. New York, NY: Oxford University Press.
- Shiraki, T., Kojima, D., & Fukada, Y. (2010). Light-induced body color change in developing zebrafish. Photochemical & Photobiological Sciences, 9(11), 1498. doi: 10.1039/c0pp00199f
- Simões, M., Breitkreuz, L., Alvarado, M., Baca, S., Cooper, J. C., Heins, L., … Lieberman, B. S. (2016). The Evolving Theory of Evolutionary Radiations. Trends in Ecology & Evolution, 31(1), 27–34. doi: 10.1016/j.tree.2015.10.007
- Snyder, H. K., Maia, R., D’Alba, L., Shultz, A. J., Rowe, K. M. C., Rowe, K. C., & Shawkey, M. D. (2012). Iridescent colour production in hairs of blind golden moles (Chrysochloridae). Biology Letters, 8(3), 393–396. doi: 10.1098/rsbl.2011.1168
- Springer, M. S., & Murphy, W. J. (2007). Mammalian evolution and biomedicine: new views from phylogeny. Biological Reviews, 82(3), 375–392. doi: 10.1111/j.1469-185x.2007.00016.x
- Van der Kooi, C. J., Stavenga, D. G., Arikawa, K., Belušič, G., & Kelber, A. (2020). Evolution of Insect Color Vision: From Spectral Sensitivity to Visual Ecology. Annual Review of Entomology, 66(1). doi: 10.1146/annurev-ento-061720-071644
- Wakefield, M. J., Anderson, M., Chang, E., Wei, K. J., Kaul, R., Graves, J. A. M., Grützner, F., & Deeb, S. S. (2008). Cone visual pigments of monotremes: Filling the phylogenetic gap. Visual Neuroscience, 25(3), 257–264. doi: 10.1017/S0952523808080255
- Wallace A. R. (1889). Darwinism: an exposition of the theory of natural selection with some of its applications. London & New York: Macmillan.
- Walls, G. L. (2016). The Vertebrate Eye and Its Adaptive Radiation (Classic Reprint). Fb&c Limited.
- Yokoyama, S. (2000). Molecular evolution of vertebrate visual pigments. Progress in Retinal and Eye Research, 19(4), 385–419. doi: 10.1016/S1350-9462(00)00002-1
- Young, J. Z., & Nixon, M. (1991). The Life of Vertebrates (3rd ed.). Oxford University Press.