
- 本文轉載自特有生物研究保育中心,《自然保育季刊》第 116 期
- 作者/鄭錫奇|行政院農業委員會特有生物研究保育中心研究員兼主任秘書、林清隆|社團法人台灣蝙蝠學會秘書長、林融|社團法人台灣蝙蝠學會研究專員、許家維|行政院農業委員會特有生物研究保育中心計畫助理、張簡琳玟|行政院農業委員會特有生物研究保育中心助理研究員
探知瞭解臺灣狐蝠 (Pteropus dasymallus formosus) 在臺灣過去的歷史及目前的現況,對於保育這種瀕臨絕種保育類野生動物至關重要。首先,我們想要從人類還沒登上綠島定居的時期談起。
據說綠島是臺灣狐蝠的故鄉,當時生存在島上的族群可能有 1,00-2,000 隻之多。原始綠島雖然森林茂盛、食物資源豐富,然而面積有限的自然環境所能涵養的狐蝠族群數量可能在千隻之譜,所以有些年輕個體可能因為競爭壓力下向外播遷,以尋求更大的生存機會。

綠島距離最近的臺東僅約 33 km,對於這種具飛行能力的大型哺乳類而言,就近遷移到臺東地區根本不是問題,並順勢逐漸擴及花蓮亦屬合理,所以歷史文獻告訴我們,花蓮老早就有狐蝠的分布;近期的研究亦證實花蓮的狐蝠和綠島的個體親緣關係密切。數百年來,原住民居住在綠島上,並利用島嶼及海域的自然資源謀生;之後,漢人亦陸續登島移居。然而,可怕的事情約莫發生在半個世紀前,當人類大舉開發綠島天然棲地並大肆捕捉利用野生動物起,臺灣狐蝠逐漸沒有了明天⋯⋯
臺灣狐蝠的發現及身世
臺灣狐蝠是臺灣最大型的蝙蝠,模式標本的來源是當時一位駐臺的英國長老教會牧師 Hugh Ritchie 自打狗 (今高雄) 獲得一對狐蝠雌雄成體標本寄給倫敦動物學會 (Zoological Society of London) (現存於英國博物館),這是臺灣狐蝠首次發現的紀錄。後經動物學家 Philip Lutley Sclater 在 1873 年發表命名為臺灣特有種 Pteropus formosus (Sclater 1873),距今已近 150 年;雖然論文中並無明確的形態描述,僅認為此種外形很像分布於琉球的琉球狐蝠(P. dasymallus),但附有J. G. Keulemans 手繪的逼真彩圖 (Sclater 1873)。1931 年堀川安市在《台灣哺乳動物圖說》寫道:「狐蝠,台灣特產,所知產地在台東廳下的火燒島(綠島),但最近在花蓮港街附近亦有捕獲。」可見當時花蓮地區就有狐蝠出沒。1933 年黑田長禮認為臺灣狐蝠與琉球狐蝠極為相似,遂修訂臺灣狐蝠的分類地位為琉球狐蝠的 5 個亞種之一,學名改為 P. d. formosus,其原產地應在綠島,而在臺灣地區所發現的其他零星個體皆屬於意外飄泊者 (Kuroda 1933)。
根據林良恭教授 (1983) 早年的論文指出,臺灣狐蝠過去的分布地點,為高雄,花蓮,綠島。至於蘭嶼,早先似乎被認為理所當然應有狐蝠存在,然而鹿野忠雄 (1929) 發現過去一直以為棲息在島中洞穴的狐蝠,其實是白腰雨燕 (Apus pacificus);不過黑田長禮 (1938) 則確認高橋定衛曾於 1933 年 10 月在蘭嶼獲得狐蝠標本。此外,1940 年黑田在《原色日本哺乳類圖說》 一書論及花蓮的哺乳類時指出:「在花蓮的哺乳類共有 10 種。……狐蝠,加禮宛……。」表示當時在花蓮地區的加禮宛有狐蝠分布。加禮宛原為噶瑪蘭平埔族遷移至花蓮地區所設七社之一,光復後改為花蓮縣新城鄉嘉里村。

幾近絕種的飛行哺乳類
臺灣為保育野生動物資源,在 1989 年訂定頒布《野生動物保育法》,保育類野生動物名錄中將臺灣狐蝠列為瀕臨絕種保育類野生動物。其實臺灣在光復之後,一直到 80 年代對於野生動物族群與分布現況的瞭解都極其有限。王穎教授在 1986 至 1988 年曾針對當時在臺灣頗為盛行的山產店對野生動物利用的情形進行一系列的調查及訪談,結果發現,在 78 家山產店中間商規模的實際調查量和一年交易推估量的前 5 名分別為山羌 (Muntiacus reevesi micrurus)、野兔 (Lepus sinensis formosus)、白鼻心 (Paguma larvata taivana)、山羊 (Capricornis swinhoei) 和野豬 (Sus scrofa taivanus),數量較少的末 5 名為水鹿 (Rusa unicolor swinhoei )、石虎 (Prionailurus bengalensis chinensis)、黑熊 (Ursus thibetanus formosanus)、黃喉貂 (Martes flavigula chrysospila) 和水獺 (Lutra lutra chinensis);至於狐蝠的交易資料完全沒有紀錄。
堀川安市在 1925 年曾在臺東的街上購買一隻狐蝠飼養,並清楚記載著飼養當時的情形 (堀川 1925)。本文第一作者曾於 1990 年在臺東地區昔日的小野柳山產店發現一隻關在籠子兜售的狐蝠,業者表示這一隻狐蝠是來自綠島。林良恭與裴家騏 (1999) 透過訪談得知,早期在綠島的獵人會連結數張尼龍製鳥網,設置於稜線上,一夜最多可捕捉 30 隻左右的狐蝠,捕獲後輾轉販售至臺灣花蓮和臺東等地,再轉手至各動物園或由私人飼養。李玲玲和林良恭於 1992 年撰文探討臺灣地區中大型哺乳動物的現況時,即認為臺灣狐蝠已幾近絕種,與雲豹(Neofelis nebulosa brachyura)和水獺同樣面臨極大的生存危機,其存續狀況令人擔心。

臺灣狐蝠的原鄉
位於臺東外海的綠島,原名火燒島,島嶼面積僅 15.1 km2,為臺灣第四大附屬島嶼,距離臺東最短的直線距離僅約 33 km。早先有達悟族等原住民登島居住,後來陸續有漢人移居;在臺灣戒嚴時期曾經是關押政治犯的著名監獄所在,如今則是以觀光產業聞名,來自國內外登島欣賞風景、體驗海洋生態與潛水活動的遊客絡驛不絕。文獻指出,在日治時期有多位日籍學者論及綠島的臺灣狐蝠,並咸認為綠島是臺灣狐蝠的產地 (林良恭 1983;吳永華 2004)。最早是岸田久吉 (1924) 在東京帝國大學發現一個可能是在 1897 年春天採自火燒島、不甚完整的臺灣狐蝠標本;黑田長禮 (1925) 提及於 1911 年在火燒島採獲的臺灣狐蝠;堀川安市 (1925) 指出,產於火燒島的臺灣狐蝠,白天隱身於岩洞 (作者註:此應是當時的誤判),天黑才出現,嗜食果實,以榕樹的果實為主食;鹿野忠雄 (1929) 亦敘述火燒島的臺灣狐蝠棲息於密林中,倒掛於樹枝,多風的日子很容易捕獲。

然而,一般民眾若在今日登島觀光旅遊,想要目睹空中飛行或棲息林間的狐蝠,機會微乎其微。林良恭及裴家騏曾於 1993 至 1996 年於綠島進行為期二年半的實地調查,但並未發現狐蝠,僅根據訪問當地居民指出,60-70 年代時期狐蝠最高族群量曾達 2,000 餘隻,然而在 1976-1986 年間遭受大量獵捕及棲地林相改變而導致族群量銳減 (林良恭 與裴家騏 1999)。綠島曾棲息 2,000 隻狐蝠,到底可信度如何?
林良恭與裴家騏 (1999) 依據 Wiles et al. (1989) 針對馬里亞納群島的瑪麗安娜狐蝠 (Pteropus mariannus) 所建立的島嶼面積與狐蝠族群量之公式推算,當未遭受到嚴重狩獵,綠島面積可供養 813 隻狐蝠,然而若有嚴重狩獵的情況下,則僅存 21 隻狐蝠。近年來,我們在綠島進行現況調查時也對當地耆老進行訪談,耆老提到有一位專門獵捕狐蝠的獵人,在一生當中就曾在綠島打到幾百隻、甚至上千隻的狐蝠。
近年綠島狐蝠調查
關於狐蝠在綠島被發現的報導資訊並不多,大概僅有 1991 年綠島居民為防止野鳥啄食木瓜園,架設鳥網意外捕獲一隻狐蝠,以及 1995 年顏聖紘教授在綠島進行昆蟲調查時於龜灣目擊過狐蝠 (吳慧雯 2010)。直至 2005 年,行政院農業委員會特有生物研究保育中心(以下簡稱特生中心)及台灣蝙蝠學會的調查團隊為執行「臺灣地區野生動物多樣性資源之調查研究」計畫,曾於當年 4 月登島進行為期一週的調查,結果僅發現幾隻零星的臺灣狐蝠 (鄭錫奇等 2006)。臺北市立動物園研究團隊於 2005 至 2009 年在綠島曾進行數年調查,亦發現過幾隻倖存的臺灣狐蝠,並於 2009 年 4 月間觀察到一隻母蝠抱著幼蝠活動,認為族群可能尚有增長潛力 (吳慧雯 2010; 陳湘繁等 2009),當時推估綠島狐蝠僅約 12 隻。
在《野生動物保育法》公告實施 27 年之後,特生中心與林務局於 2016 年根據 IUCN 的評估標準進行臺灣陸域哺乳動物之保育等級評估,並編撰出版《2017 年臺灣陸域哺乳類紅皮書名錄」(鄭錫奇等 2017),此名錄將臺灣狐蝠列為國家極度瀕危 (NCR, National critical endangered),與歐亞水獺同一等級。近年 來政府為回應社會大眾對瀕危野生動物積極進行保 育或復育的期待,在林務局統籌規劃的「國土生態保育綠色網絡建置計畫」項下,特生中心及台灣蝙蝠學會於 2018 至 2021 年合作執行「臺灣狐蝠研究與保育策略研擬案」,結果在綠島一些特定棲地陸續發現狐蝠的食渣和排遺,並在黃昏或清晨以定點觀察方式,多次發現狐蝠飛行個體,最後根據島上發現的狐蝠食渣數量推算,綠島約有 20 隻狐蝠 (範圍 10-31隻),認為目前綠島的狐蝠族群應處於數量不多但穩定的狀態 (林清隆等 2020;鄭錫奇等 2021)。

龜山島狐蝠相見歡
位於宜蘭外海的龜山島因外形似浮龜而得名,頭城人常稱之為龜島或龜嶼,面積僅約 2.84 km2, 直線距離宜蘭的烏石港僅約 10 km。昔日的龜山島有居民居住,直到 1977 年成為軍事管制區後,島上居民全數遷移至宜蘭頭城。1999 年政府將龜山島納為東北角暨宜蘭海岸國家風景區內,依當地的人文特色及生態資源,規劃為海上生態公園,並結合鄰近海域的賞鯨活動,成為北部藍色公路熱門的旅遊景點;然為減低觀光旅遊對環境造成衝擊,乃管制遊客數量並禁止在島上過夜。開放登島觀光期間偶有聽聞有人在島上發現狐蝠,但一直沒有正式紀錄。直到 2006 至 2009 年間,臺北市立動物園研究團隊登島進行調查時不僅確實觀察到狐蝠的蹤影,也發現狐蝠在島上終年活動,並有繁殖育幼行為 (陳湘繁及吳慧雯 2010);然而訪談之前的居民表示昔日在島上不曾目擊過狐蝠。
2009 至 2013 年,陳湘繁教授持續針對龜山島臺灣狐蝠族群進行自然史、棲地利用與族群遺傳結構研究,並在 2010 年根據捕捉標放個體推算島上的臺灣狐蝠族群至少有 20 隻 (陳湘繁及李涵君 2014)。至此龜山島確認為是臺灣狐蝠一處重要的棲地,然而陳教授推論,龜山島上的狐蝠族群應是晚近年代才經由海洋播遷移入,最有可能是來自八重山群島的狐蝠,因為親緣研究證據顯示,相對於與其他的臺灣狐蝠族群(如綠島),島上的狐蝠與來自八重山群島西表島 (Iriomote) 個體間親緣關係相近。2018-2021年,特
生中心及台灣蝙蝠學會團隊執行「臺灣狐蝠研究與保育策略研擬案」之歷年調查都在島上發現比陳教授當年更多的狐蝠食渣和排遺,也經常在清昏或夜間觀察到狐蝠,甚至在 4‒8 月間偶可發現母蝠帶領著幼蝠或亞成蝠活動的景象。根據狐蝠食渣的數量、目擊狐蝠個體數,以及年度間參與繁殖育幼之成幼蝠數量綜合推算,龜山島應存在有 80 多隻狐蝠(範圍 40-128 隻)。這些年來,龜山島上狐蝠族群量的整體趨勢似乎是向上的(林清隆等 2020;鄭錫奇等 2021)。

被遺忘的花蓮狐蝠
雖然如前所言,臺灣狐蝠分布於花蓮地區在日治時代就有文獻紀載 (Yasuichi 1931;Kuroda 1940),但是似乎沒受到太多的關注,近年來在花蓮發現狐蝠竟然是 50 多年後的 1992 年。1992 年,廖美菊老師記憶當時任教於花蓮女中時,曾在校園中看過狐蝠(廖美菊 私人提供)。
之後,在 1995 年至晚近幾年則陸續有人在花蓮市美崙山、美崙溪畔及出海口,市區的花蓮高農、花蓮酒廠文創園區與學校校園、鄰近的鳳林鎮發現狐蝠。2018‒2021 年,特生中心與台灣蝙蝠學會執行「臺灣狐蝠研究與保育策略研擬」時,除了研究人員每季的實地調查外,並廣邀當地的自然觀察者擔任狐蝠監測志工,從 2020 年 9 月迄今的志工登錄人數已有 38 人,並陸續回報監測資料超過 300 筆,這對於花蓮地區狐蝠的分布、活動和族群量的瞭解貢獻良多。更難能可貴的是,這些愛鄉惜蝠的監測志工不僅發現多處臺灣狐蝠在花蓮市區的日棲所 (day roost),也觀察到以往不曾知道的資訊,諸如冬季當地的狐蝠會取食王棕 (Roystonea regia,即大王椰子) 的花穗。
花蓮地區的狐蝠不僅終年都在花蓮市區活動,而且和上述 2 個孤懸外海的離島不同,牠們幾乎就生活在人們的週遭,棲息或覓食在美崙山的林間、隱身在校園公園的大王椰子叢中、流連於美崙溪畔結實累累的樹上⋯⋯,更重要的是,曾有志工在 4 月間觀察到母蝠背負著仔蝠在市區的樹上活動的景象。毫無疑問地,臺灣狐蝠在花蓮地區持續存在著一穩定族群,且是臺灣本島唯一且重要的分布區域。

我們初步推算花蓮地區應存在著約 40 隻狐蝠(範圍 29‒52 隻)。然而,花蓮的狐蝠族群從何而來?根據陳湘繁教授的族群遺傳結構研究,花蓮的狐蝠的親緣關係與綠島個體相近,而與龜山島的族群較遠 (Chen et al. 2020),或許透露一些端倪。
臺灣狐蝠都吃些什麼?
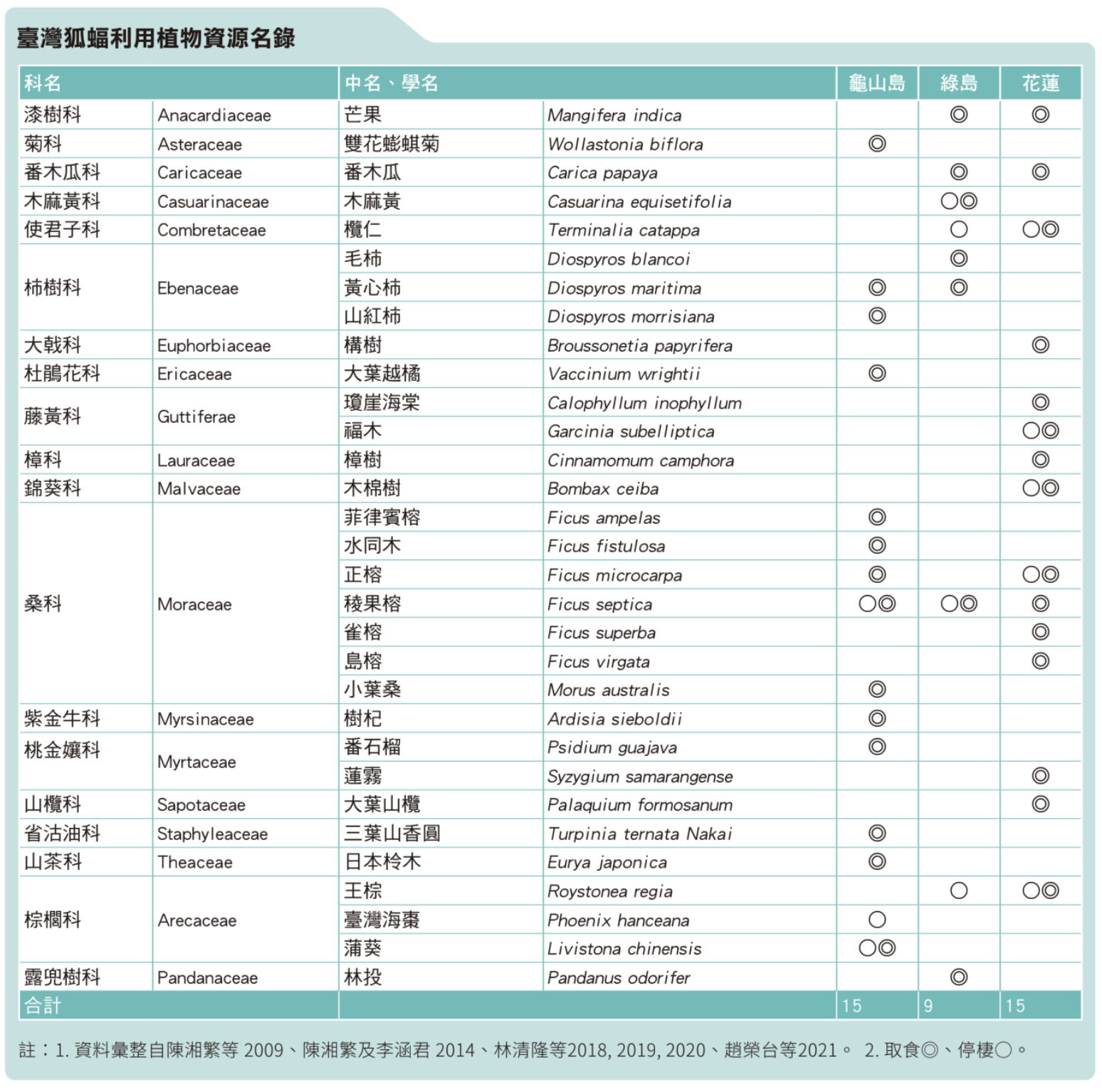
民以食為天,野生動物亦然;在今日臺灣已無濫捕野生動物的情事發生下,棲地品質與食物資源就成為族群存續相當重要的因素。我們調查發現龜山島、綠島及花蓮的狐蝠族群取食的類別有所不同,端視當地具有的植物種類,以及季節性開花結果之物候情形而定;不過桑科 (Moraceae) 榕屬 (Ficus) 的物種還是最多,這也是狐蝠最主要的食物來源,尤其是稜果榕。
臺灣狐蝠是標準的素食者,喜食植物的果實或花粉,取食果實時通常會在嘴巴內細細咀嚼、吸食汁液,最後將含有纖維的殘渣吐出,稱為食渣;狐蝠的食渣為乾扁狀並帶有齒痕,可與其他共域的哺乳動物 (如松鼠) 分辨。陳湘繁及李涵君 (2014) 在龜山島調查臺灣狐蝠的食性,發現其主要以稜果榕為食,其他尚包括水同木、正榕、雀榕、菲律賓榕、樹杞、三葉山香圓、小葉桑、日本柃木、番石榴、黃心柿等物種的果實。我們根據文獻紀錄及近年在龜山島、綠島及花蓮地區三處臺灣狐蝠主要棲息地之實地調查發現,牠們會取食或利用的植物至少有 30 種,除了上述種類外,其他尚包括芒果、番木瓜、欖仁、毛柿、山紅柿、構樹、大葉越橘、瓊崖海棠、福木、樟樹、蓮霧、大葉山欖、林投等植物的果實,以及欖仁樹葉、王棕和蒲葵的花穗、木棉樹和雙花蟛蜞菊的花朵、木麻黃的花粉等 (陳湘繁等 2009;林清隆等 2018, 2019, 2020;趙榮台等 2021)。
至於當地許多高大的樹種則往往成為牠們重要的隱棲處所。另外,調查期間我們在花蓮地區發現一個有趣的現象,即搜尋撿拾的狐蝠食渣中至少有 5 顆含有金龜子科 (Scarabaeidae) 昆蟲的碎片 (林清隆等 2020),而陳湘繁教授在龜山島連續長達 4 年的調查亦發現有 2 顆含有金龜子科昆蟲碎片的食渣 (Chen et al. 2017);巧的是,這些食渣都是在當年 7 月間所發現的紀錄。這真是非常特別而稀罕的資料,狐蝠會不會偶爾也想要開個葷呢?狐蝠的主食榕果常存在著共生的榕小蜂 (如 Blastophaga verticillata),所以當狐蝠在大啖榕果時,有可能會將小蜂或其他附著在榕果上的昆蟲給吃了下去;還是在特定的季節,狐蝠會想來點昆蟲補充蛋白質?原因真耐人尋味。

保育臺灣狐蝠的重要性
狐蝠是翼手目 (Chiroptera) 狐蝠科 (Pteropodidae) 狐蝠屬 (Pteropus) 的大型蝙蝠,現生的物種約有 65 種,主要分布在舊世界的熱帶、亞熱帶大陸與海洋島嶼 (Hall and Richards 2000)。牠們因鼻吻突出似狐狸而被稱為狐蝠 (fox bat),以花朵、花粉、花蜜、果實與樹葉為主食,因此亦被稱為果蝠 (fruit bat)。
狐蝠 (果蝠) 因具有長距離的飛行能力,以及扮演著植物授粉及種子傳播的重要角色,被視為熱帶和亞熱帶地區 (含島嶼) 維持與拓殖森林生態系的基石物種 (keystone species) (Wilson 2002)。根據研究,全世界植食性蝙蝠取食 28 目 67 科約 530 種被子植物,並協助其授粉結實及傳播種子 (Fleming et al. 2009),而被狐蝠吃下肚所排出的種籽,其發芽率顯著提高、發芽所需時間相對縮短。Fujita and Tuttle (1991)研究發現,舊世界熱帶植物有 289 種需要蝙蝠協助授粉和傳播種子,總計可以產出 448 種通稱為蝙蝠產物 (bat-dependent products) 的經濟物品,諸如食物、藥材、木材、染料、燃料、飲料原料、水果、纖維、飾物、以及其他多項的森林副產品。然而,許多研究報告也指出,近數十年來由於颱風或極端氣候劇變的影響,以及人類開發、變更林相或直接獵捕所造成的嚴重結果,常導致狐蝠族群數量急劇下降,尤其以海島族群為甚 (Allison et al. 2008;Chaiyes et al. 2017; Esselstyn et al. 2006;Mickleburgh et al. 2008; Nakamoto et al. 2011;Struebig et al. 2007; Welbergen et al. 2008)。

根據現況調查的結果推估,臺灣地區的狐蝠族群尚不及 200 隻,避免滅絕的族群數量恐嚴重不足!那到底需要多少族群數量,才足以讓狐蝠永久存續在臺灣地區?根據 Lin 等人 (2021) 最近對臺灣狐蝠遺傳多樣性的研究結果發現,由遺傳結構推估,臺灣狐蝠由 2,324 隻的歷史族群數量因重大影響而衰退至目前的 223 隻,而且其族群遺傳變異度偏低,並有相對高的近親交流指數。因此,我們認為綠島原來擁有的族群數量或可當作參考值,也就是說 1,000‒2,000 隻之數應是臺灣狐蝠可以永續的族群量。
Chen 等人 (2020) 新近的研究指出,出現在龜山島的狐蝠族群呈現較高程度的遺傳歧異度,除了自己獨特的支系外,也有部分個體與臺灣本島、以及鄰近的八重山狐蝠有較近的親緣關係,而其高遺傳歧異度 (相對於綠島的族群)導因於多個遺傳分群混合的結果,因此推測龜山島族群可能包含多個祖先的起源,不同時期陸續有個體移入補充;論文最後並建議,孤立島嶼的亞種族群應該分開管理,並需努力減少其遺傳多樣性的持續下降。
臺灣狐蝠目前主要分布在綠島、龜山島和花蓮三個區域,各地的族群所面臨的生存威脅不盡相同,在保育作為和經營管理上必須要有整體思維及地域上的策略。我們在執行「臺灣狐蝠研究與保育策略研擬案」計畫,最終也提出一份《臺灣狐蝠保育行動計畫書》,除了列出「臺灣狐蝠族群擁有可存續的族群量以達到止跌、回升、脫離困境」及「讓民眾能正確認識臺灣狐蝠並友善對待,不再有不當利用與誤解」的二項計畫願景外,也根據地區性的生存威脅因子提出短、中、長期的保育執行目標與策略,期盼經由多方的努力,儘快降低臺灣狐蝠在臺灣地區滅絕的機率。瀕危物種的保育或復育刻不容緩,而維繫一群健康且永續的狐蝠族群,不論對牠們生存的生態系或是對人類的福祉而言,都相當重要。































-200x200.jpg)









{kind=link}