- 本文轉載自 自然保育季刊 111 期《太田樹蛙的重新命名─修正電子期刊所衍生不適用的分類作為》
- 作者 / 王盈涵(國立臺灣大學生態演化所碩士)、蕭郁薇(國立臺灣師範大學生命科學系碩士)、李閣桓(澳洲麥覺理大學博士生)、曾惠芸(國立臺灣大學昆蟲學系助理教授)、林彥博(行政院農業委員會特有生物研究保育中心助理研究員)、小卷翔平(日本岩手醫科大學助理教授)、林思民(國立臺灣師範大學生命科學院教授)
最近這 3 年,臺灣兩棲爬行動物最大的變革, 大概就是「 日本樹蛙」(Buergeria japonica) ( 圖 1 ) 在分類地位上的修正。 過去所認定的日本樹蛙分布廣泛, 從臺灣往東北至琉球群島北端, 甚至跨越了重要的生物地理學屏障—吐噶喇海峽 (Tokara Gap), 而分布到吐噶喇群島 (Tokara Islands)。這麼廣泛的幅員,自然也成為生物地理學家探討物種分布時重要的研究題材。從 1990 年代中期,當時任教於琉球大學的太田英利 (Hidetoshi Ota) 教授、國立自然科學博物館周文豪 (Wen-Hao Chou) 研究員、京都大學松井正文 (Masafumi Matsui) 教授等人,就已經針對日本樹蛙的族群差異進行初步的分析,顯示在整個物種之中存在幾個遺傳上高度分化的類群 (Tominaga et al. 2015)。但由於這些不同地點的日本樹蛙外觀非常近似,叫聲又細碎龐雜,很難從單一的形態或聲音特徵區分牠們的差別。
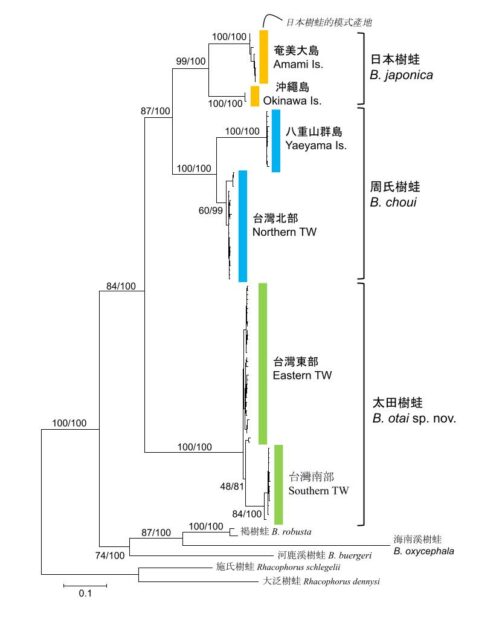
圖 1:溪樹蛙屬 (Buergeria) 的親緣關係。這棵演化樹是使用粒線體 DNA 細胞色素b (cytochrome b) 序列,採用最大似然法 (maximum likelihood) 所建構。/林思民 製圖。
直到 2017 年,國立臺灣師範大學生命科學系的研究團隊,利用遺傳、形態、聲音、行為四組證據, 將臺灣東部與南部的族群發表為獨立的新種太田樹蛙 (B. otai) (Komaki et al. 2017;Wang et al. 2017) (圖 2)。

日本琉球大學與京都大學的團隊,再利用遺傳與形態的差異,將八重山群島與臺灣西北部的族群稱為周氏樹蛙 (B. choui) (Matsui andTominaga 2020)。這兩個物種的名字分別向太田英利教授和甫從科博館退休的周文豪副館長致敬,以感謝他們對東亞島弧兩爬研究的貢獻。
但是分類學上的問題並未因此落幕!2017 年秋,在太田樹蛙的論文刊登的隔週,我們收到國外友人的來信,信中善意的提醒:我們發表的期刊 PLOS ONE 是電子期刊,並未刊行紙本,這導致了命名過程的瑕疵。按照過往國際動物命名規約 (International Code of Zoological Nomenclature, ICZN, https://www.iczn.org/) 的規定,新種的命名必須刊行在紙本的期刊上,才算完成正式的「發表」。但是在這個資訊爆炸的網路時代,圖書館已經逐漸取消蒐藏紙本論文;而許多國際期刊也逐漸朝向無紙化的電子期刊邁進。為了因應時代的改變,國際動物命名規約提供了一個解決方案:只要事先在「動物學名銀行」 (ZooBank) 上註冊,宣告命名新種的作為,最後在電子期刊上提供相關的註冊編號,國際動物命名規約就可以承認這個物種的正式發表 (第三章 8.5 條)。
閱信之後心想,我們真的遭遇了一個難以解決的窘境!每個科學家的論文在被接受之前,都必須經過冗長的投稿和退稿過程。我們投稿的論文被前兩份排行更高的紙本期刊婉拒之後,嘗試的第三份期刊就是 PLOS ONE。雖然我們並不是第一次發表新種,也並不是第一次投稿電子期刊,但是這兩個元素的致命組合,就在我們的投稿過程發生了。經過提醒,我們趕緊在 “ZooBank” 上註冊了太田樹蛙,同時去函給 PLOS ONE 的編輯部。依從編輯部的建議,我們在 2018 年 2 月發表了一個補正文件 (correction) 做為補救,把 2017 年未及時附上的註冊編號收錄在文件之中 (Wang et al. 2018)。國際上數個全球性的兩棲類名錄網站,隨即也將太田樹蛙收錄其中,並將新種有效的時間從 2018 年 2 月起算。
然而一波未平,一波又起。正當我們鬆一口氣的時候,馬上又收到幾位分類學者的善意來函:我們做的彌補仍然不完備!這其中又包含了幾派不同的解讀。首先,在正式的命名規約之中,新種的命名必須要伴隨著模式標本的指定、保存、存放地點和鑑別特徵等說明(第四章 16.4 條)。雖然我們 2017 年的論文已經對模式標本進行詳細而完整的描述,但是在 2018 年的補正文獻中,我們沒有再次提及這些重覆的資料。如果把 2017 年的論文主體和 2018 年的論文補正當作是「同一次」的分類作為,兩份文件合起來確實是齊全的。但是如果有些分類學家從嚴認定,把 2017 年和 2018 年當做是「兩次」不同的命名作為,那麼第一次缺了 “ZooBank” 序號,第二次缺了模式標本的詳細說明,兩次的作為都不合法。另有一派的說法則認為:既然國際動物命名規約認為 “ZooBank ID” 必須要在「事前」註冊,那不管再怎麼彌補,這件事情都已經難以回溯。最好乾脆從頭再起爐灶,也就是說,再進行一次新種的描述。

看到這邊,相信諸位讀者已經跟我們一樣,被國際動物命名規約的繁瑣規定沖昏了頭。規約的存在是為了維持學名系統的穩定性,畢竟每一個新種都是分類學家曠時費力的心血結晶,必須受到規約的嚴格保護。但是當碰到新世代的電子期刊制度和百家爭鳴的網路媒體時代,更必須特別注意其中的細節,以避免造成分類上的混亂。雖然這個案例中同時存在著模稜兩可的不同觀點,但我們體會到:當分類學上發生爭議的時候,從嚴認定才是最好的解方。所以,我們勢必得進行第三次的分類作為。

其中一個方法就是請 PLOS ONE 再出版第二次補正,但是我們仍然沒有把握這個作為是否能成立;第二個釜底抽薪的方法,則是乾脆再重新命名這個新種。如果這次的命名符合所有國際動物命名規約的規定,爾後提及太田樹蛙時,牠引述的文獻就將由本文取代。更多的詳細資料,包含遺傳、形態、叫聲、行為等等資訊,既然在 2017 年的文章之中都已經闡述,那些內容我們就不再重複;但是新種的命名作為,就讓這篇文章重新定義。
以下,我們就宣告命名這個新種。
樹蛙科 Family Rhacophoridae Hoffman, 1932(1858)
溪樹蛙屬 Genus Buergeria Tschudi, 1838
太田樹蛙 新種 (Buergeria otai sp. nov.)
正模標本:
NMNS 19819 ( 圖 3 ),成年雄性個體。採自臺灣屏東縣東港溪流域 (北緯 22.626340,東經 120.643342),2015 年 7 月 14 日由王盈涵、呂嘉偉採集。 以 10% 福馬林固定後移入 75% 酒精,存放於國立自然科學博物館。這隻個體的鳴叫聲亦可在 Wang et al. (2017) 的線上資料中下載。
副模標本:
包含 22 隻雄性成蛙與 5 隻雌性成蛙。雄性樣本: NMNS 19815、19817、19822、19824 採集自屏東縣東港溪;NMNS 19825、19831、19833採集自屏東縣楓港溪;NMNS 19856、19859、19865 採集自花蓮縣瑞穗鄉;NMNS 19846、19850、19853 採集自花蓮縣八里灣溪;NMNS 19788、19799、19803 採集自花蓮縣美崙溪;NMNS 19806、19808、19812 採集自宜蘭縣南澳溪;NMNS 19838、19840、19844 採集自宜蘭縣蘭陽溪。雌性樣本:NMNS 19867、19868、19869 採集自花蓮縣美崙溪;NMNS 19870、19871 採集自宜蘭縣南澳溪。由王盈涵、李閣桓、林展蔚、呂嘉偉、曾文宣、蕭郁薇、蔡緯毅、李宜龍、林雨昕、沈敬家、陳濠森、林品萱等人共同採集。以 10% 福馬林固定後移入75%酒精,存放於國立自然科學博物館。
命名依據:
本物種的學名 otai 是將太田英利教授(Hidetoshi Ota)的姓氏拉丁化,以表彰太田教授對臺灣以及東亞兩棲爬行動物的研究貢獻。我們建議使用的中文名為「太田樹蛙」,亦可以稱為「太田氏樹蛙」或「太田氏溪樹蛙」。
鑑別特徵:
太田樹蛙是小型的樹蛙科成員,身體纖瘦。雄性成蛙的體長 23.1 ‒ 29.3 mm (平均值 26.57,測量樣本 133 隻);雌性成蛙的體長 29.7 ‒ 37.5 mm (平均值 32.44,測量樣本 3 隻)。背面有輕微的不規則顆粒狀突起,其中在肩胛骨之間的一對短棒狀突起最為明顯。指端與趾端膨大,形成吸盤。前足指尖無蹼,後足趾間部分有蹼。肩胛骨至背部通常有一個 X 形或 H 形的暗色花紋,下頷與腹部灰白色,四肢有深棕色的寬橫紋。大腿腹側有密集規則的小白點,通常集中於大腿基部。
近緣物種的比較:
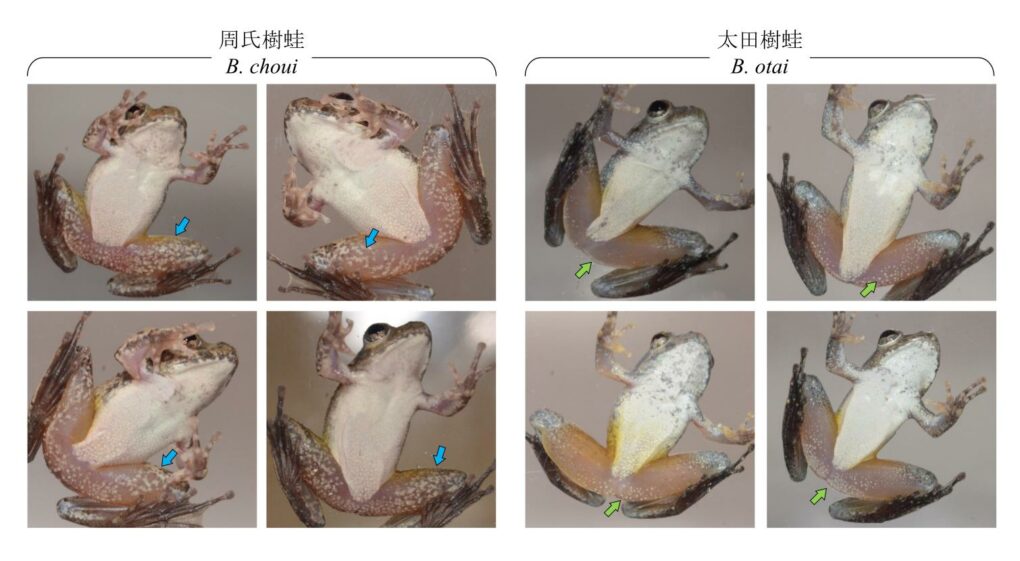
形態上與太田樹蛙最接近的類群是臺灣北部與南琉球群島的周氏樹蛙,以及琉球群島北部的日本樹蛙。與這兩種近緣樹蛙相比,太田樹蛙的頭部在統計上明顯較大,但是肉眼不易區分。因此,可靠的鑑別方式來自大腿基部的斑點與叫聲訊號。當平貼在透明玻璃或壓克力表面時,可見太田樹蛙的大腿腹側呈現許多大小與形狀規則的小白點。雖然個體之間的白點數目差距甚大,但是原則上會集中在大腿的基部。相較之下,周氏樹蛙的大腿腹側呈現的是不規則的雲狀斑,與太田樹蛙的斑點有明顯的差別 (圖 4)。
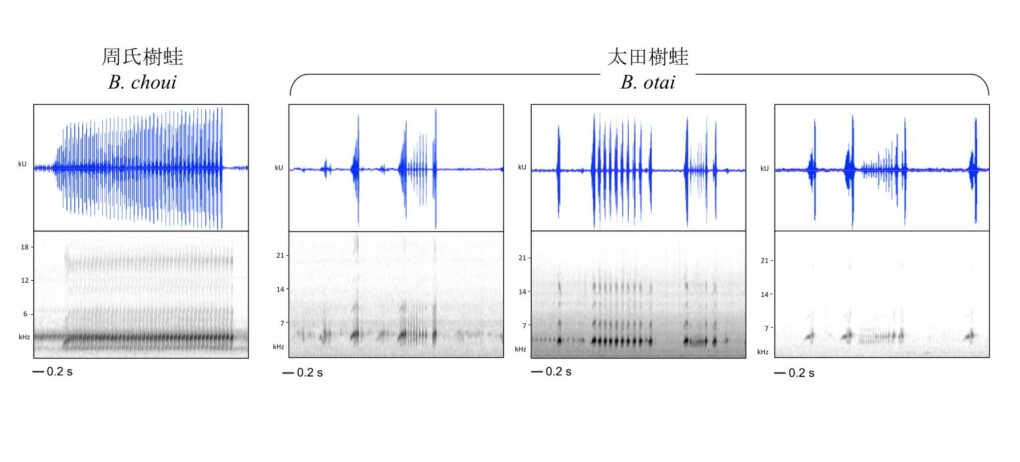
在聲音訊息上,太田樹蛙、周氏樹蛙及日本樹蛙這三者都會發出定位用的短鳴叫和宣示用的長鳴叫,也導致過去在鑑別上的挑戰。實際上周氏樹蛙和日本樹蛙都只會發一種長鳴聲,這種長鳴聲的速度和強弱都很均勻,乍聽之下類似細碎的蟲鳴 (圖 4)。太田樹蛙則會發出兩種長鳴叫,其中第一種叫聲類似周氏樹蛙與日本樹蛙,但是叫聲的節奏較緩慢。第二種叫聲則富含節奏變化與強弱起伏,經常由一個高強度的單音起始,經過細碎的呢喃之後,又以高強度的單音作收。由於周氏樹蛙不會發出第二種叫聲,因此如果在溪流中聽到忽強忽弱的鳴唱聲,當地的族群即可鑑定為太田樹蛙 (圖 4)。
結語
自從 2017 年首次描述之後,利用次世代定序技術結合基因體的分析,加上行為測試的樣本增加,我們發現太田樹蛙和周氏樹蛙在叫聲上的差異確實會影響到牠們的擇偶條件,進而影響遺傳上的區隔。這些證據證明太田樹蛙和周氏樹蛙確實存在明顯的遺傳與行為分化,而且有許多與性擇相關的行為議題,均值得深入進行研究。然而,依循著過去兩百多年的傳統,從公開的發表 (publication),到成為一個適用名稱 (available), 進而讓牠在分類上正式而有效 (valid),這仍然是物種分類學必須依循的傳統流程。所幸在我們補救的過程中,獲得科博館鄭明倫研究員和海生館何宣慶研究員的大力釜正。我們也感謝多位來自全球的分類學家,包括世界兩爬學界的兩位泰斗 David Wake、松井正文 (Masafumi Matsui),以及Dr. Darrel Frost、富永篤 (Atsushi Tominaga)、江頭幸士郎 (Koshiro Eto)、徐堉峰、顏聖紘等多位熟悉命名規約的教授與先進,先後提供我們寶貴的意見,並耐心等候我們完成正規的命名處理。文章獲得自然保育季刊審查委員詳盡的修改意見,也謹此一併致謝。
太田樹蛙在分類作為上的一波三折,其實是時代交替的過程之中好幾項錯誤同時發生的結果。但既然身為作者,我們就得承認錯誤發生的事實,進而設法改正。除了帶給大家一則茶餘飯後的八卦故事,或者成為分類學課堂上老教授講古時的怪奇案例,我們更希望太田樹蛙曲折離奇的命名過程可以讓大家學會教訓,不要再走上我們歷經的冤枉路。
近年來,分類學受到大眾的忽視,而得到的經費和資源挹注也逐年遞減;但是其中仍然有著博大精深的內涵蘊含其中,絕對不可等閒視之。
- 詳細論文修正內容請參考:自然保育季刊 111 期《太田樹蛙的重新命名─修正電子期刊所衍生不適用的分類作為》原文附錄。