演化歷史是如何被改寫的?基於形態證據的系統發育分析或許能為我們提供更多證據,來揭示 2 億 2600 萬年前的化石真正的「歷史」。
現今的分類學家將鞘翅目 (Coleoptera) 分為四大亞目,牠們分別為原鞘亞目 (Archostemata)、藻食亞目(Myxophaga)、肉食亞目 (Adephaga)、以及多食亞目 (Polyphaga)。
而其中隱翅蟲科 (Staphylinidae)為多食亞目中物種數最多的一群,同時牠們甚至是動物界中已描述物種數最豐富的一群。隱翅蟲科可說是演化最成功的一類生物,因此牠們的演化歷史一直是昆蟲學家著迷的課題。
想了解更多有關隱翅蟲的資訊,可見本站延伸閱讀:

L. prorova 是最古老的隱翅蟲?其實牠是藻食亞目啦
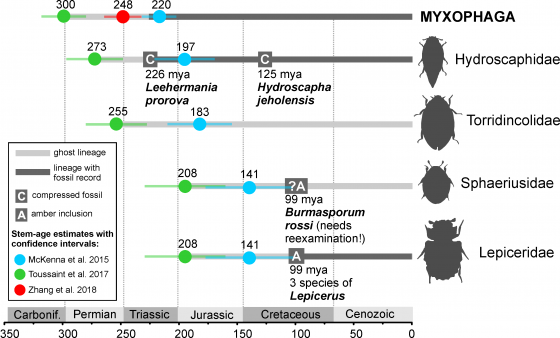
以往被認為是最古老的隱翅蟲是三疊紀的化石物種 Leehermania prorova (Chatzimanolis, Grimaldi& Engel, 2012) 雖說在發表之初就有被懷疑可能不是隱翅蟲,但一直到最近才正式被揭示了,L. prorova 應屬於藻食亞目的成員。該研究由捷克國家博物館的昆蟲學家 Martin Fikáček 與各國的昆蟲學家組成的團隊完成,分析了其與現生甲蟲科別的系統發育關係,相關成果最近刊登於頂尖昆蟲學期刊《系統昆蟲學》(Systematic Entomology)。
如何分析化石與其他現生甲蟲的系統發育關係?
研究團隊結合現生甲蟲所有科別的形態特徵矩陣(包括化石)建構出各種不同的系統發育樹,他們使用最大簡約法 (Maximum parsimony) 及貝葉斯推斷 (Bayesian Inference) 分別分析所有甲蟲科別、隱翅蟲總科(Staphylinoide)、藻食亞目及 L. prorova 的資訊建構系統發育樹。所有證據都支持 L. prorova 不是隱翅蟲,研究團隊最後提出 L. prorova 是已滅絕的藻食亞目支系,最接近的現生科別為出尾水蟲科(Hydroscaphidae),同時,L. prorova 也成為最古老的藻食亞目成員。

太小啦,大顎還是小顎鬚傻傻的分不清?
雖然受限於 L. prorova 體型非常小(僅約2 mm),有非常多的特徵無法檢視,但研究團隊亦將牠與隱翅蟲比較後,提出幾個特徵從而推斷牠並非隱翅蟲。其中最有趣的莫過於在 L. prorova 的原始描述中提及牠有大顎,但研究團隊發現那其實並不是大顎,而是有分節的小顎鬚 (Maxillary palpi),這個發現也支持牠不是隱翅蟲,因為隱翅蟲的大顎通常於背面觀或側面觀清楚可見,而藻食亞目則幾乎不可見。

當今化石系統分類研究的典範
此一研究除了確認最老的隱翅蟲並不是隱翅蟲這個重要的發現,同時它也樹立了當代化石系統分類文章的典範。分子鐘的應用,在當今的系統分類研究中是很常使用的工具。而化石證據能夠校準分子鐘,除了使用現生物種對比化石外,此研究也不止使用傳統的形態特徵矩陣,亦參考分子資料來佐證形態證據,將所有的證據同時檢視,而提出最有可能的研究結果。相信在不久的將來會有越來越多研究能使用此方法來推斷生物的演化歷史。
參考文獻
- Fikáček, M., Beutel, R. G., Cai, C., Lawrence, J. F., Newon, A., Solodovnikov, A., Ślipiński, A., Thayer, M. K. & Yamamoto, S. 2019. Reliable placement of beetle fossils via phylogenetic analyses–Triassic Leehermania as a case study (Staphylinidae or Myxophaga?). Systematic Entomology.
特別感謝 Martin Fikáček(捷克國家博物館)提供圖片及蔡晨阳(中國科學院南京地質古生物研究所)協助校稿。