公元 1823 年華勒斯在英國誕生,於 1913 年以 90 歲高壽去世,今年 2023 年是他誕生 200 年。我們懷念他是因為,他曾經和達爾文聯名發表演化觀點,以及提出解釋東南亞海島間生物分佈差異的「華勒斯線」。
Alfred Russel Wallace 在台灣常翻譯為華萊士,不過如威廉華勒斯等等 Wallace 都翻譯作華勒斯,本文就統一作華勒斯。
達爾文會提出演化論,深受他知名的小獵犬號之行影響。華勒斯的東南亞考察也給予他不少啟示,一如達爾文的加拉巴哥群島等地;然而在此之前,華勒斯已經在亞馬遜有 4 年經歷。為了紀念華勒斯兩百歲生辰,Nature 期刊 2023 年初刊登兩篇文章,緬懷他的亞馬遜之旅。
 華勒斯 1860 年代的畫像,當時 40 歲左右。圖/Mondadori Portfolio via Getty
華勒斯 1860 年代的畫像,當時 40 歲左右。圖/Mondadori Portfolio via Getty
與強者朋友一起前進亞馬遜,然後分道揚鑣
和前輩達爾文相比,華勒斯的家境普通,也沒有受過正規的學術研究訓練。所幸身處文化發達的大英帝國,有志青年仍有不少學習和出人頭地的機會。何況他爸爸是學過法律的自耕農,文化資本其實不算低。
成年後喜歡生物的華勒斯在 1844 年,21 歲之際遇見小他 2 歲的貝茲(Henry Walter Bates),兩人志同道合;華勒斯也從一般愛好者,升級為有系統的標本搜集者,可謂一隻腳踏入研究領域的門檻。
1848 年,華勒斯 25 歲之際與貝茲一同航向大西洋對岸的亞馬遜。不過兩人大部分時候分開行動,貝茲在亞馬遜南部,華勒斯在北部的尼格羅河(Rio Negro)一帶。
華勒斯年輕時在談笑無鴻儒,往來皆白丁的階段,我猜朋友大概不會只有貝茲一位。不過貝茲後來提出的貝氏擬態(Batesian mimicry)沿用至今,可謂華勒斯的強者我朋友,事後諸葛的我們建構歷史敘事時,也就津津樂道兩人的友誼。
英國病人碰上船難,買保險很重要!
離家萬里的華勒斯,依然透過經紀人與國內保持聯繫,郵寄異鄉產品回英國賺錢。在亞馬遜待了 4 年後他決定返鄉,期間一直被疾病威脅生命,可謂現實意義上的英國病人(The English Patient)。
最慘的是他弟弟 1849 年遠渡重洋來照顧他,卻自己也感染黃熱病,返國途中不幸病逝。而華勒斯要等到幾個月後才收到消息。
1852 年華勒斯搭乘海倫號(Helen)貨船返國,沒想到出海三個星期後火燒船,使他漂浮在大西洋海面上,眼睜睜看著攜帶的行李大多損毀。最後他耗費 80 天返回英國,比起與貝茲的去程 29 天漫長得多。好在經紀人有買保險,讓華勒斯獲得部分補償,不至於血本無歸。
 返回英國的海倫號火燒船事件後,沒有損毀的少數紀錄。圖/The Natural History Museum/Alamy
返回英國的海倫號火燒船事件後,沒有損毀的少數紀錄。圖/The Natural History Museum/Alamy
回到英國的華勒斯將近 30 歲,闖出一些名號,卻沒有受到太多重視。所幸保住生命加上幾年累積的知識,賦予他東山再起的契機。1854 年他得到前往東南亞的機會,1858 年 35 歲時就和達爾文聯名發表歷史巨作。
從亞馬遜參透生命的奧秘:生物地理學
華勒斯僅管在亞馬遜一直生病,也淬煉出不侷限於觀察的科學眼光,從船難撿回一條命回到英國後,展露學術鋒芒。1852 年 12 月 14 日,他在倫敦的動物學會發表研究亞馬遜猴子的論文,主張亞馬遜各地的猴子款式,受到大河形成的地理障礙強烈影響。
當時華勒斯受到一些批判,後來證明他的論點無誤,而且是生態分佈的普遍現象。現在我們知道更多:亞馬遜的河道歷史上改道多次,導致生物的分佈範圍持續變化。
用現代標準看,前往亞馬遜考察的 4 年差不多等同華勒斯的博士班修行,回國後發表的報告則是他的博士論文。這篇博士級論文中還觸及一個要點,所謂的「亞馬遜雨林」內部其實差異不小,他是首先有意識提及此事的研究者。
華勒斯觀察到亞馬遜的不同地區,物種組成不太一樣。他劃分 4 大區域:幾內亞、厄瓜多、秘魯、巴西,由其間的亞馬遜河、尼格羅河、馬德拉河(Madeira)這些大河分割出不同地區的地理障礙。如今所知更多,還可以切得更細。
具體是觀察到有幾條河分割出幾塊地,超乎其上普世性的生物學道理是,由於地理環境的阻隔,各地會形成不同的「特有種(endemism)」。華勒斯領悟地理障礙會影響生物分佈,可謂生物地理學的先驅。
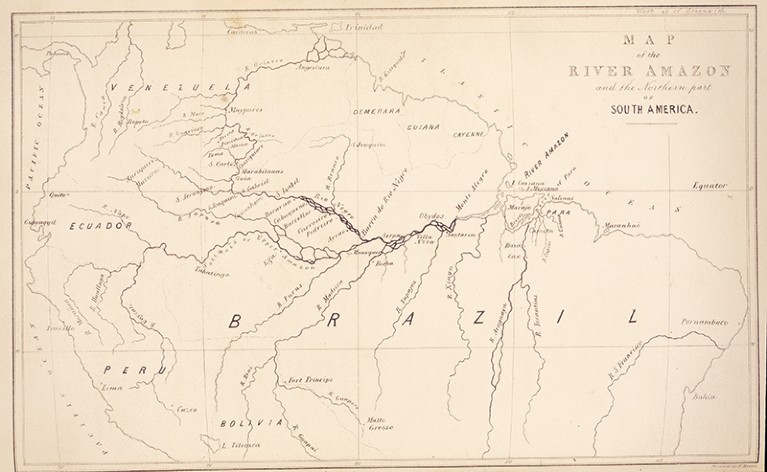
 華勒斯 1853 年出版書籍中的亞馬遜地圖。圖/Mary Evans/Natural History Museum
華勒斯 1853 年出版書籍中的亞馬遜地圖。圖/Mary Evans/Natural History Museum
自學成才的英國洞觀者
現在的人可能覺得上述觀點都是些普通常識。可是華勒斯是在 1852 年提出,那時演化論尚未問世,跟他同齡的孟德爾,當時也尚未開始種植豌豆。
一百多年後的常識,首度問世時常常是驚天動地的新突破!
年輕的華勒斯沒有受過正規學術訓練,還是需要持續賣標本換錢的月光族,提出的研究成果竟有如此理論性。由此可知亞馬遜之行,確實讓華勒斯從所謂的集郵者,蛻變為具備洞察力的科學家。
法國詩人韓波(Arthur Rimbaud)認為,詩人必需是能看穿事物表面,有洞察力的洞觀者(voyant),我想這也是頂級科學家必需配備的能力,亞馬遜的神秘力量加持過後,華勒斯可謂成功通靈。
這類自學成才的科學家,當時英國不只華勒斯一位。以時代來說,那時的英國社會有點厲害。後來華勒斯沈迷於「唯靈論(spiritualism )」就是另一個故事了……
華勒斯年輕的南美洲經歷,讓人聯想到更早將近一百年的洪堡(活到很老,1859 年 90 歲時去世)。身為晚輩,華勒斯讀過洪堡作品,他站在洪堡巨人的肩上,觸及到更高的思想境界。
許多人覺得遺憾,遺傳、演化並稱,但是孟德爾提出遺傳學法則後被埋沒超過 30 年,等到 1900 年代才重現於世,因此 1882 年去世的達爾文沒有機會知悉。這方面華勒斯比較幸運,他年紀比孟德爾小半歲,又一直活到 90 歲,有機會見證遺傳學的發揚光大。
 華勒斯 1853 年出版書籍中提到的「黑暗中一團燦爛之火(sitting amidst the gloom, shining out like a mass of brilliant flame)」圭亞那動冠傘鳥。圖/Hein Nouwens/Getty
華勒斯 1853 年出版書籍中提到的「黑暗中一團燦爛之火(sitting amidst the gloom, shining out like a mass of brilliant flame)」圭亞那動冠傘鳥。圖/Hein Nouwens/Getty
燦爛之火多年以後依舊燃燒
多年在亞馬遜、東南亞走跳的華勒斯,有不少接觸原住民的機會。照文字紀錄看來,他年輕時的思想應該和同時期的普通英國人差距不大,沒有特別進步或反動;不過相比於同時代人,他更尊重在地知識,這也有助於他的成功。
亞馬遜的生物多樣性如今依然天下第一,世界卻變化不少。尼格羅河盆地的原住民,在華勒斯時代是被觀察者,類似實驗動物的角色,現在漸漸變成主動的研究者,他們用源自不同文化的手法探索自己的世界,成為現代知識體系的一份子。
然而,曾經啟發華勒斯的尼格羅河盆地,至今仍缺乏一流的研究機構,無法培育本土的研究人才,本地學子必需離鄉背井。科學從華勒斯到現代突飛猛進,仍有不少進步空間。
上圖是華勒斯描述為「黑暗中一團燦爛之火」的圭亞那動冠傘鳥(Guianan cock-of-the-rock ,學名 Rupicola rupicola),目前沒有滅團危機,依然在華勒斯探索過的雨林中飛翔。希望燦爛之火永不熄滅,但是不要變成失控的森林大火。
延伸閱讀
參考資料
- Alfred Russel Wallace’s first expedition ended in flames
- Escaping Darwin’s shadow: how Alfred Russel Wallace inspires Indigenous researchers
- Evolution’s red-hot radical
本文亦刊載於作者部落格《盲眼的尼安德塔石匠》暨其 facebook 同名專頁。