美國國家海洋和大氣管理局(The National Oceanic and Atmospheric Administration, NOAA)組成一支調查團隊,USGS的國際野生動物保育中心的野生動物專家,利用專業儀器,將病毒從海豹的身體組織中分離出來,專家們指出其中有一型禽流感病毒,與野生鳥類的H3N8禽流感病毒十分接近。
但當時第一次世界大戰的主要參戰國家,如德、英、法、美等國為了避免影響士氣,嚴格管制媒體報導疫情。然而保持中立而未參戰的西班牙,因為沒有實施戰時審查制度,西班牙媒體自由報導著流感相關新聞,甚至連西班牙國王阿方索十三世(King Alfonso XIII)感染重症的消息也被廣泛報導,造成西班牙疫情特別嚴重的錯覺,也因此被命名為「西班牙流感」。
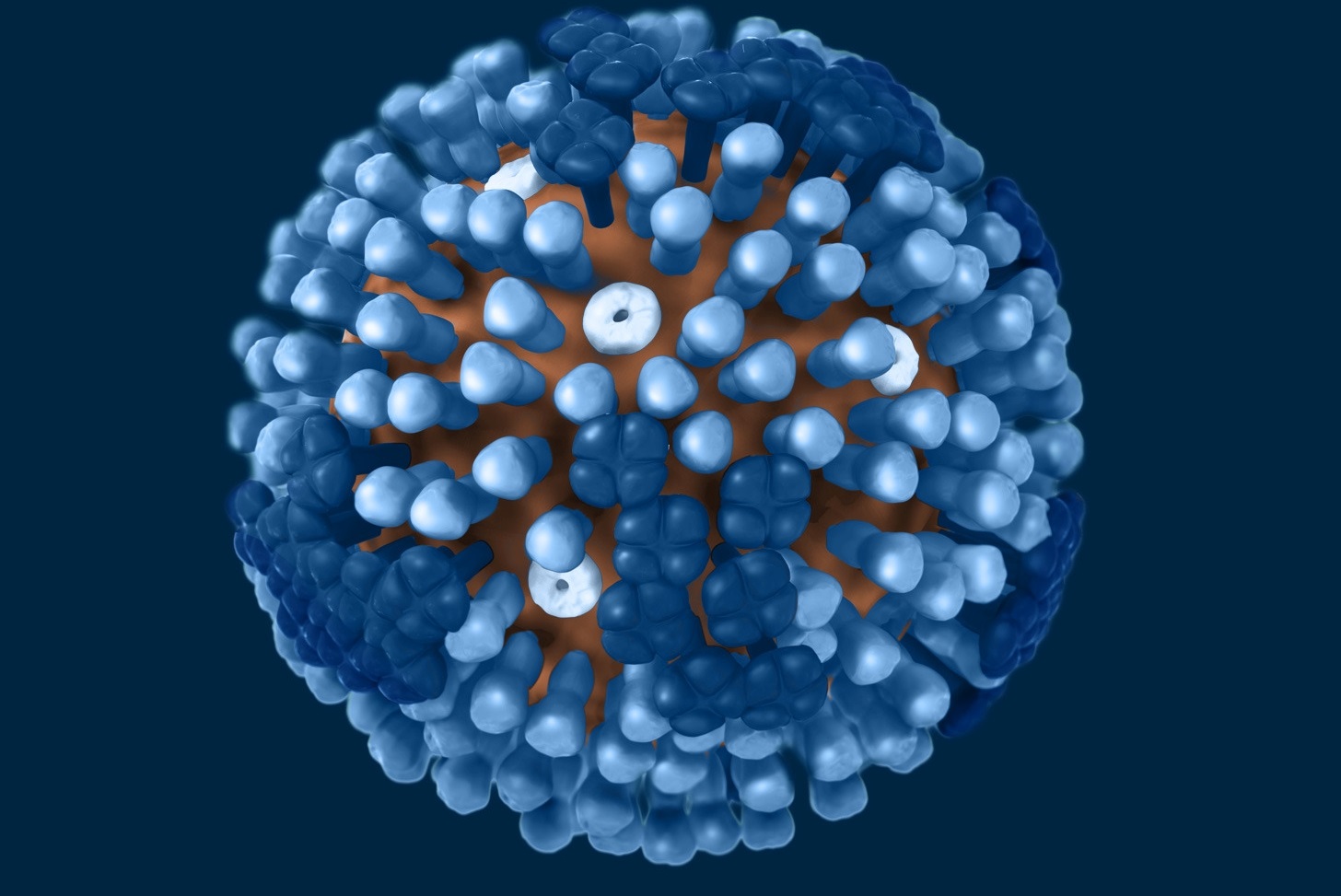
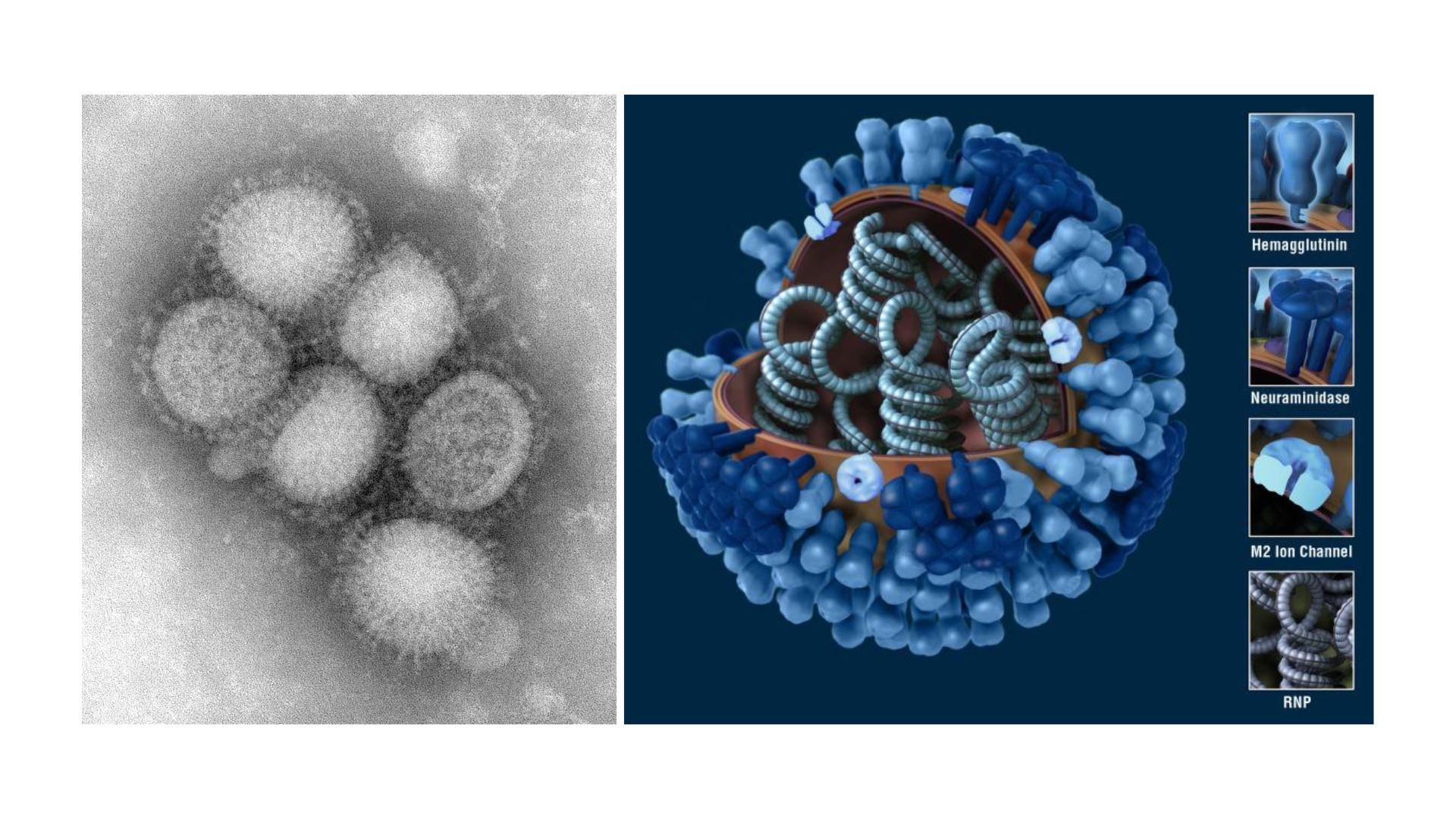
病毒外殼上還零星分布M2離子通道蛋白(M2 ion channel protein),但數量非常少,平均每100至200個HA,才有一個M2。套膜下則有M1基質蛋白(matrix protein M1)支撐病毒結構,維持病毒穩定。B型流感病毒的整體結構和A型非常類似,只是膜蛋白組成略有不同,除了HA和NA之外,另有兩種B型流感獨有的NB和BM2蛋白。至於C型流感病毒,外型就和A、B型明顯不同,它們在感染細胞表面時,能形成長達數百微米的「繩索狀結構」。
Bouvier, N. M., & Palese, P. (2008). The biology of influenza viruses. Vaccine, 26 Suppl 4(Suppl 4), D49–D53.
Berche P. (2022). The Spanish flu. Presse medicale (Paris, France : 1983), 51(3), 104127.
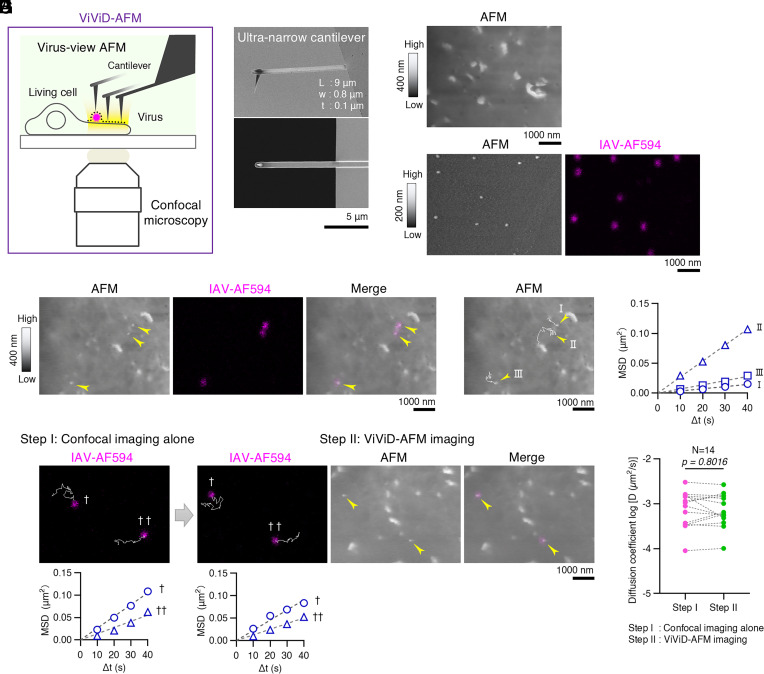
Yoshida, A., Uekusa, Y., Suzuki, T., Bauer, M., Sakai, N., & Yamauchi, Y. (2025). Enhanced visualization of influenza A virus entry into living cells using virus-view atomic force microscopy. Proceedings of the National Academy of Sciences of the United States of America, 122(38), e2500660122.











































{kind=link}