最近,英國牛津(Oxford)大學的 Jamie Warner 研究團隊與日本電子公司(Japan Electron Optics Laboratory)合作發現一個能減少色差效應的新方法。他們使電子探測樣本前先經過一單色儀,此步驟會縮減電子能量範圍並因而有效增進空間解析度。解析度的提升使得該團隊能首次以真實原子級解析度來研究石墨烯內的刃差排(edge dislocation)。刃差排是材料內一種特別的缺陷形式,會造成晶格結構的扭曲。
赫爾穆特.魯斯卡(Helmut Ruska,約 1969 年)由艾德曼.魯斯卡(Erdman A Ruska)提供。圖片來源:〈Helmut Ruska and the visualisation of viruses〉
人類對微觀世界的探索,最早可以追溯到17世紀。當時,英國博物學家羅伯特‧虎克(Robert Hooke)利用自製顯微鏡觀察軟木塞,觀察到了植物細胞壁,並稱其為「細胞」(cell)。荷蘭的雷文霍克(Antonie van Leeuwenhoek)以精湛的磨鏡技術,進一步製造出放大倍率更高的顯微鏡,在清澈的水中發現了肉眼見不到的「生物」,成為第一個發現細菌、紅血球和精子的人。
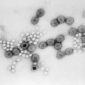
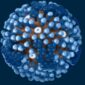
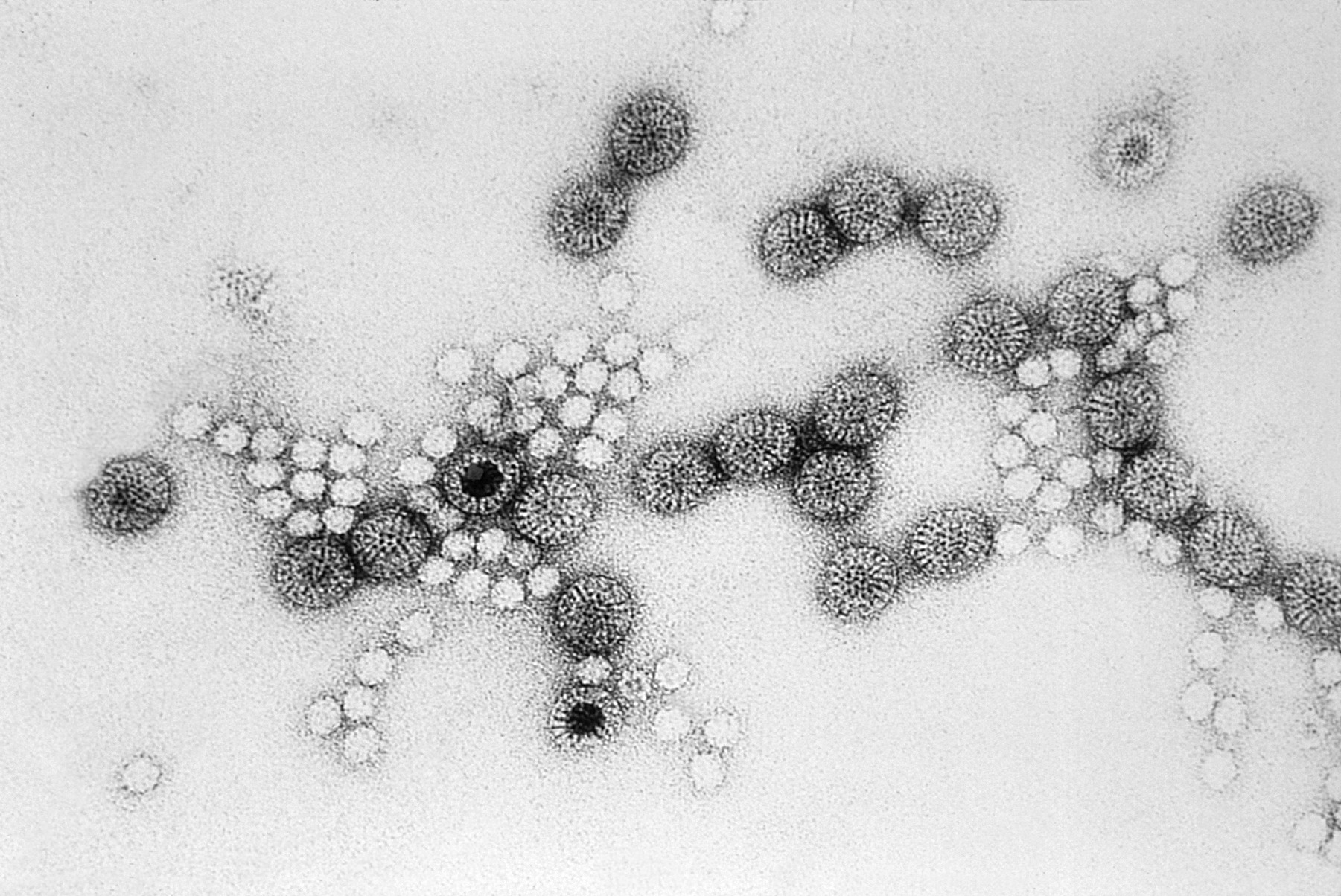
1940年代初,赫爾穆特已發表了約20篇關於細菌、寄生蟲和不同病毒超顯微結構的報告,這些出版物標誌著首次利用電子顯微鏡對病毒進行視覺化。包括1939年他與考舍(Gustav A. Kausche)和普凡庫赫(Edgar Pfankuch)合著的《超顯微鏡下植物病毒的影像》,展示了菸草花葉病毒的桿狀結構,首次揭示病毒的亞微觀顆粒。


")