


編按:《人類這個不良品》羅列人體的各種缺陷與設計不良之處,但其實這就是演化自然產生的結果。這些不完美成就了獨一無二的我們,也讓我們照見演化的歷史。
一條條軸突傳遞神經訊號
人類神經系統的複雜性和重要性簡直到了驚人的地步。我們有高度發展的腦子,而腦子得透過神經才能發揮功用。
軸突就像一條一條獨立的細小纜線,負責在腦部和身體各處之間來回傳遞神經脈衝,聚集成束的軸突就是神經。好比位於腦部頂端的某些運動神經元就有極長的軸突,這些軸突延伸至腦部以外, 沿著脊髓下行,離開腰椎區,再沿著雙腿往下,最後抵達大腳趾。
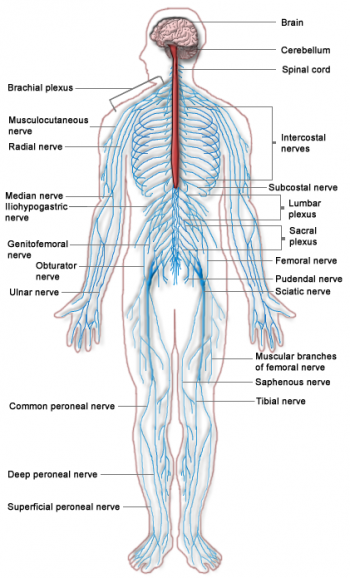
這條路徑雖然漫長,但目的直接又明確。腦神經和脊神經的軸突則有如一張綿密的網,從腦部出發,分布至人體各個肌肉、腺體和器官。
經過大動脈再回到喉頭,繞遠路的喉返神經
在人類的神經系統裡,演化作用同樣留下了古怪的缺失。就拿「喉返神經」(recurrent laryngeal nerve,簡稱RLN)來舉例,先容我在此說明:人體多數神經是成對存在的,左右半身各一條,不過為求敘述方便,姑且就以左半身的喉返神經為例。
喉返神經的軸突從腦部頂端附近起源,並與喉頭的肌肉相連。喉頭肌肉受到神經的指揮,讓我們在說話、悶哼和唱歌時,能夠發出聲音,並加以控制。
始於腦部,終於喉嚨上半部,這條路徑理應很短:經過脊髓, 進入喉嚨,抵達喉頭,大不了幾公分的距離吧?
錯了。喉返神經的軸突包覆在一條更出名的神經—迷走神經(vagus nerve)之內。迷走神經自脊髓往下抵達上胸部,自此喉返神經才從迷走神經中分支出來,從肩胛骨稍下處離開脊髓,接著, 左喉返神經繞經大動脈下方,然後再重新回到頸部,抵達喉頭。
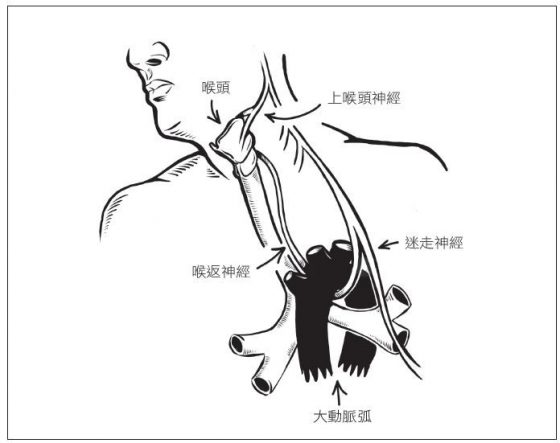
喉返神經的總長度足足比預定長度多了三倍以上,繞經不需經過的肌肉和組織,和許多心臟大血管互相交纏,是心臟外科醫師替病人手術時,得特別小心注意的一條神經。
都是演化搞的鬼?喉返神經要追溯至古老的魚類
早在古希臘時代,著名的加倫醫生就發現這個古怪之處。如此迂迴的神經行進路線,有什麼功能上的意義嗎?幾乎沒有。事實上,同為支配喉頭肌肉的上喉頭神經(superior laryngeal nerve),行進的路線就完全切中我們的預期。上喉頭神經同樣由更大的迷走神經中分支出來,在腦幹下方就離開脊髓,然後直接抵達喉頭,既簡單又明瞭。
那麼,為什麼喉返神經要選擇這麼一條孤單而漫長的道路呢?答案同樣藏在古老的演化歷史裡。
喉返神經源自於古老的魚類身上,所有現代脊椎動物身上都有這條神經。魚類的喉返神經連接腦和鰓,鰓可謂喉頭的祖先。然而,魚類腦子小,沒有頸部,沒有肺,牠們的心臟比較像一條肌肉軟管,不像人類的心臟有如一顆幫浦。因此,魚的中央循環系統,位置幾乎就在鰓的正後方,這一點跟人類大不相同。
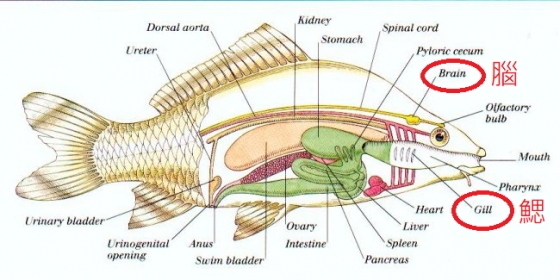
魚的喉返神經離開脊髓抵達鰓,走的是一條想當然耳又兼備效率的短路徑。魚的喉返神經在這條路途中,也確實和離開魚心的部分主要血管互相纏繞,這些血管等同哺乳類動物的動脈分支。
在魚身上,神經和血管交織的狀況是合理的,這樣才能在極度局促的空間裡,以最緊密又簡單的方式安置神經和血管。然而,隨著魚類演化出四足類,再演化出人類的過程,這樣的安置方式卻也造就了人體內荒謬的結構設計。
心臟距離腦部愈來愈遠,喉返神經只好愈來愈長
在脊椎動物演化過程中,出現了明顯的頸部和胸部,因此心臟的位置往後移動許多。從魚類到兩棲動物,兩棲動物到爬行動物,再從爬蟲類到人類,心臟的位置距離腦部愈來愈遠,然而鰓的位置並沒有變動。就解剖學的角度而言,人的喉頭之於人腦,就如同魚鰓之於魚腦。
要是喉返神經沒有和心臟血管互相纏繞,行進路線就不會受到心臟位置變動的影響。但是從腦部出發的喉返神經確實和心臟血管交纏,所以脫不了身,想要返回頸部就不得不繞這麼一大圈。顯然,想要從胚胎發育期著手,解開交纏的心臟血管和返喉神經,重新設計神經行進的路線,對演化作用而言不是一件簡單的任務。
人類的喉返神經白白繞了一大圈,經過頸部和上胸部所造成的後果,或許看起來不算太嚴重,畢竟所有四足的脊椎動物都從共同祖先「硬骨魚」那兒承襲了相同的結構設計。
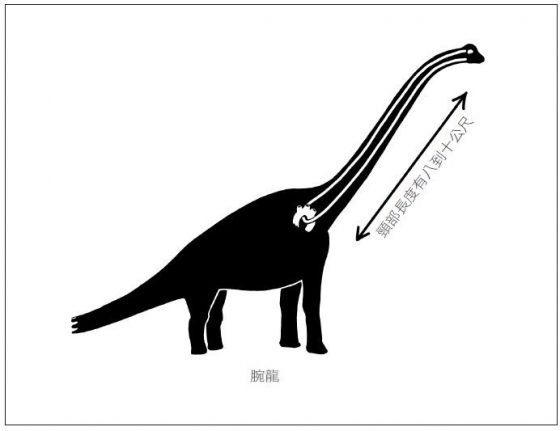
鴕鳥的喉返神經其實只需要行進二至三公分的距離,就可以發揮功用,但鴕鳥喉返神經沿脊髓下行的長度就有一公尺,再返回到頸部又是一公尺的距離。長頸鹿的喉返神經長度高達五公尺!更別提迷惑龍、腕龍,和其他隸屬蜥腳亞目的恐龍,牠們的喉返神經有多長了。這麼一比較,人類似乎應該懂得知足。

本文摘自《人類這個不良品:從沒用的骨頭到脆弱的基因》,2018 年 12 月,天下文化出版。









































{kind=link}
{kind=link}