「心酸埋於土壤,過往都遺忘,化作一顆琥珀的模樣。」《郁可唯‧琥珀》

琥珀是由古代植物分泌的樹脂經長期掩埋而最終形成的珍貴半寶石,琥珀生物化石則是在形成琥珀的樹脂在流出時「活埋」了週遭環境中的生物,是意外失足的心酸古墓,也是探索古生物的珍貴寶庫。來自緬甸克欽邦出產的琥珀年代約為 9900 萬年前 (晚白堊世、森諾曼期),保存大量完好的古代生物遺骸,為近年古生物學者所專注的其中的焦點之一。
大花蚤,寄生於昆蟲的甲蟲
大花蚤 (Ripiphoridae)是外形楔形、駝峰狀的甲蟲,幼生期時為寄生性,寄主包括蟑螂、蜂類或其他甲蟲,最古老的大花蚤化石紀錄為來自內蒙古道虎溝化石層生物群 (九龍山組,中侏儸統,約 1.65 億年前)的媧皇始源大花蚤 Archaeoripiphorus nuwa Hsiao, Yu & Deng, 2017,為筆者於今年年初發表的研究 [註1],其餘的大花蚤化石紀錄不多,大多為新生代的琥珀化石,中生代的部分則僅有的四例的種類描述自緬甸和法國琥珀。有關琥珀化石和大花蚤科的介紹也可以參考這兩篇文章〈羅塞塔始源小菊虎: 9900 萬年前失足,從此凝結在時間中〉和〈以創世女神為名的史前寄生甲蟲——媧皇始源大花蚤〉。

大花蚤目前複分成 5 個亞科,其中細身大花蚤亞科 (Pelecotominae)的翅鞘完整覆蓋住腹部,被認為是較原始的類群。目前已知細身大花蚤亞科的成員在幼生期時會寄生在朽木棲的甲蟲類群幼蟲,例如竊蠹蟲,雌蟲會將卵產在朽木上靠近竊蠹蟲幼蟲製造出的坑道開口附近,細身大花蚤的幼蟲孵化後會去尋找適合寄生的竊蠹蟲幼蟲,穿越其表皮進行內寄生,並且在隔年從寄主原先用來化蛹所製作的蛹室中破蛹而出。
目前最早的細身大花蚤亞科化石紀錄為描述自緬甸琥珀的海蒂扇角大花蚤 Flabellotoma heidiae Batelka, Prokop & Engel, 2016 [註2]。
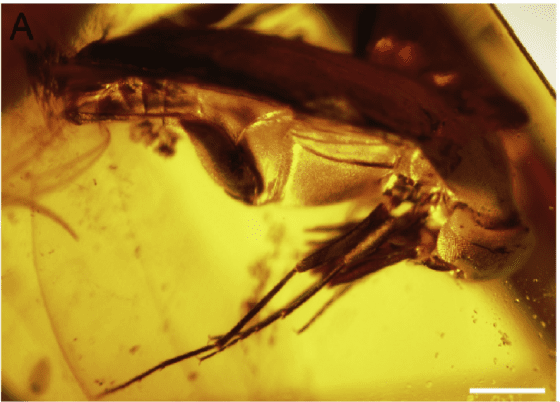
日前,同為石探記科學團隊的我和臺灣師範大學黃嘉龍博士則發現了緬甸琥珀中已知的第四種大花蚤,同時也是細身大花蚤亞科的第二筆中生代化石紀錄,研究論文發表在古生物和地質學領域國際期刊《白堊紀研究》(Cretaceous Research)。我們將其命名為:陳睿刺足大花蚤 Spinotoma ruicheni Hsiao & Huang, 2017,屬名語源來自「刺」(Spino-) 和「細身大花蚤屬的字尾」(-toma),意即其密著棘刺的足部表面;種小名則獻名給本研究的幕後功臣——陳睿博士,表彰其協助提供研究材料的貢獻,陳博士任職於中國科學院動物研究所,是研究蚜蟲生態演化的年輕學者。
這塊琥珀化石目前收藏、展示於深圳世紀琥珀博物館。

本研究發現增進了我們對史前大花蚤的生物多樣性的認識,此外透過現生大花蚤類群和已知的化石物種間的比較形態學和中生代的古環境學,我們得以對於大花蚤科形態、多樣性演化和古生物學的進行以下推論:
多樣性:
Batelka 等人在發表海蒂扇角大花蚤的研究論文中,曾提出大花蚤的物種多樣化始於中生代晚期,我們透過與早前發表的 4 種中生代大花蚤成員的形態比較,不難看出在中生代晚期,大花蚤科的確已具備相當程度的多樣性,我們的發現也印證了 Batelka 等人的論述。
形態演化:
在前人的比較形態學研究中曾提出有關大花蚤科的祖先特徵 (祖徵,Plesiomorphy) 及形態演化,認為如現生大花蚤特定類群的縮短翅鞘應是演化自完整覆蓋住腹部的類群,這個演化趨勢為目前已知的形態、分子親緣關係研究所支持,而根據我們對於已知中生代化石大花蚤的形態研究亦驗證此假說。
古生物學:
本種為細身大花蚤亞科 (Pelecotominae)的成員,已知現生的細身大花蚤亞科成員會寄生於木棲的甲蟲類群幼蟲,基於外觀形態的相似性以及緬甸琥珀生物群也出土豐富的朽木棲的甲蟲類群,則因此推測本種可能已建立與朽木棲甲蟲的寄生關係,我們並且認為這些寄生性甲蟲的多樣化可能與這些潛在的木棲寄主在白堊紀的多樣性演化相關。

侏儸五化甲的系統分類:
2011 年,中國科學院南京地質古生物研究所王博博士等人發表了一種來自侏儸紀中期的擬步行蟲總科甲蟲——侏儸五化甲 (Wuhua jurassica Wang & Zhang, 2011),研究人員認為本種為一種「似花蚤類」生物,並將其歸劃於科級未定位,而根據我們在這個研究中對其形態與先前發表的媧皇始源大花蚤和現生的鋸角大花蚤 (Trigonodera tokejii (Nomura and Nakane, 1959))比較,我們認為侏儸五化甲很可能屬於大花蚤科的成員。
花蚤類楔形體態的演化:
已出土的化石紀錄顯示,最早於侏儸紀晚期已有花蚤科甲蟲的出現,這些花蚤與現生的後裔體態十分近似,很有可能亦以花粉為食,然而牠們當時似乎不太可能以水生的古果屬 (Archaefructus)花粉為食而較有可能與裸子植物關係密切,也就是說現生花蚤體態外形並非一般所認為的是與開花植物共演化而來 [註3],基於花蚤科與大花蚤科的親緣關係以及我們的研究,我們認為現生的大花蚤科之獨特的身體結構亦非與被子植物協同演化的產物,而是直接繼承了侏儸紀時期祖先類群的身體構建。
- 此文由國立臺灣大學昆蟲學系學士蕭昀撰寫,響應 PanSci 「自己的研究自己寫」,以增進眾人對基礎科學研究的了解。
參考文獻
- Hsiao, Y., Yu, Y., Deng, C., Pang, H., 2017b. The first fossil wedge-shaped beetle (Coleoptera, Ripiphoridae) from the middle Jurassic of China. European Journal of Taxonomy 277, 1–13.
- Batelka, J., Prokop, J., Engel, M.S. 2016. New ripiphorid beetles in mid-Cretaceous amber from Myanmar (Coleoptera: Ripiphoridae): First Pelecotominae and possible Mesozoic aggregative behaviour in male Ripidiinae. Cretaceous Research 68, 70–78.
- Wang, B., Zhang, H. 2011. The oldest Tenebrionoidea (Coleoptera) from the Middle Jurassic of China. Journal of Paleontology 85(2), 266–270.
- Hsiao, Y., Huang, C.-L., 2018. Spinotoma ruicheni: A new Late Cretaceous genus and species of wedge-shaped beetle from Burmese amber (Coleoptera, Ripiphoridae, Pelecotominae), Cretaceous Research 82: 29-35. Published Online: 28 October 2017 (doi: 10.1016/j.cretres.2017.10.022).