夏天發燒喉嚨痛別硬扛!新冠流感「去季節化」反撲,醫揭3大元凶與治療黃金期


呼吸道病毒已「去季節化」,夏季也成新冠與流感高峰!醫師提醒,出現發燒、喉嚨痛切勿輕忽,應及早就醫進行多重快篩,把握3至5天的投藥黃金期,以防延誤病情引發重症。
過去大家常認為,流感或其他呼吸道感染的盛行期往往落在寒冷的冬季。然而,自COVID-19疫情爆發以來,各類呼吸道病毒的流行節奏已逐漸改變,出現了波段式起伏、季節界線模糊的現象,甚至在夏季也會發生疫情升溫的情況。新冠病毒已從早期的大規模流行,轉變為與其他呼吸道病原並存的常態化傳播型態。當「感冒只在冬天」的傳統印象被打破,民眾在出現呼吸道症狀時,該如何正確辨識風險並把握就醫時機?義大醫院感染科主任賴重旭醫師從臨床角度為我們深入解析。
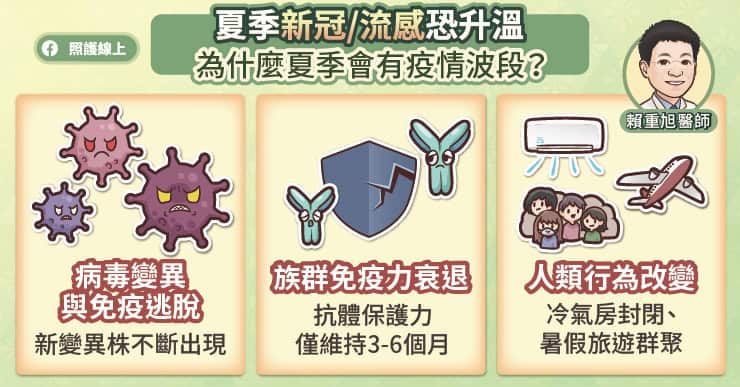
病毒去季節化!三大原因導致夏季疫情反撲
近年來,新冠、流感等病毒在夏季依然可能出現流行高峰。賴重旭醫師指出,這種「時序錯位」背後主要有三大原因:
。病毒頻繁變異:以新冠病毒為例,目前多為傳播力與免疫逃脫特性極強的衍生亞型變異株,大約每三到六個月就會產生一次變異,導致舊有的免疫防禦難以有效抵抗新病毒。
。族群保護力衰退:無論是透過自然感染或施打疫苗所產生的免疫力,保護效果通常僅能維持約三到六個月。當抗體減弱形成「免疫缺口」時,遇到新變異株就容易引發新一波感染。
。人類行為改變:炎熱的夏季促使民眾長時間待在相對擁擠且封閉的冷氣房內。此外,暑假期間的出國旅遊、跨區移動與群聚活動增加,也大幅提高了病毒傳播與互相感染的風險。
因此,現在的疫情已轉為受變異株更替與免疫力變化影響的「波段式流行」,難以再用單一季節來預測風險。
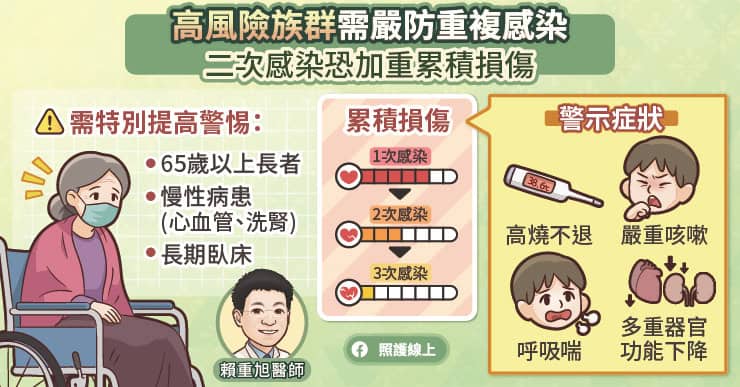
二次感染成常態!高風險族群需嚴防「累積損傷」
在波段式流行的型態下,許多民眾可能已經歷經一次甚至兩次以上的新冠感染。賴重旭醫師表示,在臨床上再次感染的狀況非常常見,甚至有些人因為症狀輕微、未加篩檢而不自知,進而無意間傳染給家中的長輩或慢性病患。
許多人誤以為再次感染的症狀一定比較輕微,但實際上不能僅以單次症狀的嚴重度來評估風險。對於65歲以上長者、患有慢性心血管疾病、洗腎病患或長期臥床等高危險族群而言,重複感染不僅可能讓身體受損的狀況慢慢累積,發展為重症或死亡的比例也明顯高於健康成年人。當高危險群出現高燒不退、嚴重咳嗽多痰、呼吸喘,甚至多重器官功能下降等危險徵兆時,務必要提高警覺。
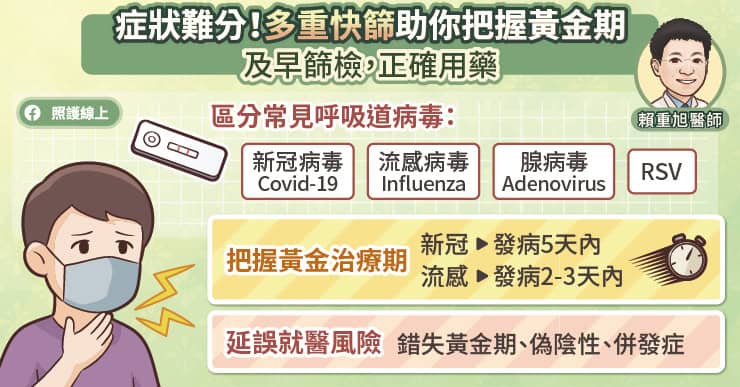
症狀相似難分辨,「多重快篩」幫助正確用藥
現今新冠、流感、腺病毒等呼吸道病毒皆可能在夏季活躍,而它們的初期表現(如發燒、喉嚨痛、肌肉痠痛等)高度相似,僅靠臨床症狀實難精準區分感染源。
為解決此困境,近年醫療院所陸續導入多重快篩工具,一次採檢就能初步辨識出多種常見病毒。賴重旭醫師強調,確認病毒種類除了能評估是否給予專屬的抗病毒藥物外,也能避免在未知狀況下使用不必要的抗生素。
賴重旭醫師也分享臨床經驗,多重快篩曾協助揪出同時感染流感與新冠的特殊案例;而在快篩全數排除常見病毒後,也曾協助醫師及時轉換感染源方向,揪出黴漿菌或披衣菌等細菌感染,進而給予正確的抗生素治療。
警惕時間差!延誤就醫恐致偽陰性與併發症
許多民眾在出現發燒或喉嚨痛時,習慣先在家觀察幾天再說,但這可能會錯失治療黃金期。賴重旭醫師進一步提醒到,常見的病毒感染:如新冠病毒的抗病毒藥物建議發病五天內投藥,而流感最好在發病兩到三天內投藥,才能達到最佳的治療效果並降低重症風險。
若拖到發病五天後才就醫篩檢,體內的病毒量可能已經降低,容易引發快篩「偽陰性」造成誤判。此外,拖延病情也可能導致單純的病毒感染惡化,併發細菌感染等其他併發症,使後續的治療方向變得更加棘手。
隨著疫情常態化,臨床醫師的評估思維也從疫情初期的「確診與隔離」,轉為重視病患的「風險分層」與病程走向。現在的門診評估會綜合考量四大面向:年齡與共病狀況、症狀惡化速度(如是否突發呼吸困難或高燒不退)、近期免疫狀況與感染次數,以及家庭同住者的風險情境。
面對氣候變遷與病毒流行的複雜化,民眾應提高警覺。當出現呼吸道症狀時,不應僅憑季節或自身感覺判斷風險,而應及早尋求專業醫師評估、妥善利用篩檢工具釐清病因。唯有把握黃金治療期,才能有效保護自己與身邊的高風險家人!
筆記重點整理
● 流感、新冠或其他呼吸道傳染病的疫情已轉為受變異株更替與免疫力變化影響的「波段式流行」,難以再用單一季節來預測風險。
● 許多人誤以為再次感染的症狀一定比較輕微,但實際上不能僅以單次症狀的嚴重度來評估風險。對於65歲以上長者、患有慢性心血管疾病、洗腎病患或長期臥床等高危險族群而言,重複感染不僅可能讓身體受損的狀況慢慢累積,發展為重症或死亡的比例也明顯高於健康成年人。
● 近年醫療院所陸續導入多重快篩工具,一次採檢就能初步辨識出多種常見病毒。確認病毒種類除了能評估是否給予專屬的抗病毒藥物外,也能避免在未知狀況下使用抗生素。
● 延誤就醫會造成更多問題,例如錯失治療黃金期,新冠病毒為發病五天內,而流感為發病兩到三天內,才能達到最佳的治療效果並降低重症風險。