- 文/王齡敏,獸醫師,社團法人台灣猛禽研究會猛禽救傷站主任。
全球各地的大城市當中,因建築的玻璃與鏡子等設計,經常會發生鳥類飛行時錯認這些是可飛行的路徑,導致撞上而造成傷亡的「窗殺」事件。窗殺可歸納為日間撞擊與夜間撞擊,兩者成因不同。
窗殺事件層出不窮,臺灣也該正視此問題,並研擬出相關的預防措施。
世界各國的窗殺事件
有一種人為的無心傷害,造成每年上億隻野鳥死亡。
這種因撞擊玻璃或建物而導致的傷害稱為窗殺 (bird-building collisions 或 bird-window collisions)。世界各地每年都有多起窗殺案例,舉例來說,美國約有 3 億 6500 萬 ~ 9 億 8800 萬起案例、加拿大約 1600 ~ 4200 萬起及南韓每年約 800 多萬隻野鳥死亡。

雖然北美對野鳥窗殺議題已研究 30 多年,但在臺灣卻很少被提及與討論,相關的研究甚至付之闕如。
回歸到窗殺的發生原因,為何鳥類特別容易撞玻璃呢?
- 首先,鳥類(或大部分的動物)無法將玻璃辨識為隔離物或障礙物;
- 第二,鳥類雖具有翅膀可飛翔並來去自如,但也容易誤判如高樓的玻璃反射影像而撞上。
北美研究窗殺議題多年,美國明尼蘇達州奧杜邦學會 (Audubon Minnesota) 於 2010 年 5 月出版的《鳥類安全之建築指南》(Bird-Safe Building Guidelines),大致將窗殺歸納為日間撞擊與夜間撞擊。
日間撞擊原因有二:
- 一為玻璃具有穿透性,因此玻璃帷幕或隔音牆等建物,若是位於鳥類可能穿越的路徑上,便會導致窗殺;
- 二為玻璃的鏡像反射效應,即使透明的玻璃也可能會產生鏡像的效果,更遑論貼有隔熱紙或特別設計為單向透視的鏡面玻璃。
甚至一些半戶外環境,如公廁或游泳池等設立的鏡子,鏡像效果導致鳥類無法分辨影像真偽,以為反射的遠景或山林影像可以飛過,因此一頭撞上,嗚呼哀哉。
夜間撞擊的原因則為燈塔效應 (beacon effect),當夜間空氣濕度較高,或有霧氣或霧霾時,建築物的燈光會吸引遷徙中的鳥類,導致鳥類迷航而誤撞建物。

明尼蘇達的窗殺案例
筆者曾於 2013~2015 年間服務於美國明尼蘇達大學猛禽中心 (The Raptor Center, University of Minnesota),當時聽聞明尼亞波利斯 (Minneapolis) 市中心將興建美國合眾銀行體育場 (U.S. Bank Stadium),並計畫採用大面積的玻璃作為建築設計。
筆者在當地工作期間曾數次路過該體育館改建前的休伯特‧漢弗萊體育場 (Hubert H. Humphrey Metrodome),由於當時對窗殺涉入不深,是無意間與一名猛禽中心的志工聊天而討論到此事,他表示很擔心這棟建築未來對於當地鳥類的衝擊。
而在筆者先前擔任野生動物獸醫師的職涯中,不時會接獲窗殺案例,但由於占傷病原因的比例並不高,所以過去筆者認為這只是不常見的偶發傷害。直到回國後,於 2017 年起在台灣猛禽研究會(以下簡稱猛禽會)進行猛禽救傷,發現為數不少的猛禽因撞窗而癱瘓,才逐漸意識到窗殺對於野鳥的衝擊。
後來明尼亞波利斯的新體育場於 2016 年落成,當地的研究人員蒐集並分析 2017 ~ 2018 年間的野鳥撞玻璃案例,他們除了合眾銀行體育場外,還監測當地其它 20 棟具窗殺風險的建築。

調查期間共蒐集 1000 多起的鳥類窗擊案例,發現合眾銀行體育場窗擊事件占所有 21 棟建築的第二高位(225 ~ 229 件),其中包括 42 種鳥類(該研究調查到的窗殺鳥種共 75 種)。
報告中指出窗殺會因季節不同而有所變化,秋季明顯高於春、夏二季(冬季因當地過去研究窗殺機率低,故此研究未將其納入),秋過境期的窗殺比率為春過境期四倍,而候鳥遭窗殺的數量則較高,前五名物種皆是候鳥,分別為白喉帶鵐 (Zonotrichia albicollis)、黃喉蟲森鶯 (Leiothlypis ruficapilla)、橙頂灶鶯 (Seiurus aurocapilla)、黃喉地鶯 (Geothlypis trichas) 與灰綠叢森鶯 (Leiothlypis peregrina),占此研究窗殺比例近 50%。
臺灣也該正視鳥類窗殺事件!
北美許多地區都有類似的研究報告與長期監測活動,但臺灣對窗殺的系統性研究目前仍未開啟,頂多只有一些零星的撞玻璃傷亡鳥類的花邊新聞報導。
筆者於去 (2019) 年起設立臉書社群「野鳥撞玻璃回報」,希望藉由網友的力量蒐集國內關於野鳥窗殺資料。另外,猛禽會也於去年獲得聯華電子主辦的「綠獎」青睞,計畫今 (2020) 年於臺灣北部地區執行野鳥窗殺調查與友善鳥類玻璃教育推廣,希望引起社會大眾、企業與政府對於野鳥窗殺的重視。
該如何避免窗殺?
看到這裡,或許讀者會急著想知道到底如何防止野鳥窗殺。其實江湖一點訣,說破不值錢,原理就在於想辦法讓鳥能「看到」或「看懂」眼前的玻璃(無論窗戶、鏡子或隔音牆等)是無法通過的阻隔物。
因此,凡是改善玻璃材質,如霧面、雕花或蝕刻圖案等;玻璃上裝飾,如貼或畫上密集圖案或大面積圖案等;與玻璃外布置,如掛上許多垂墜物、植生牆、圍欄和隱形鐵窗等,都有防治效果。
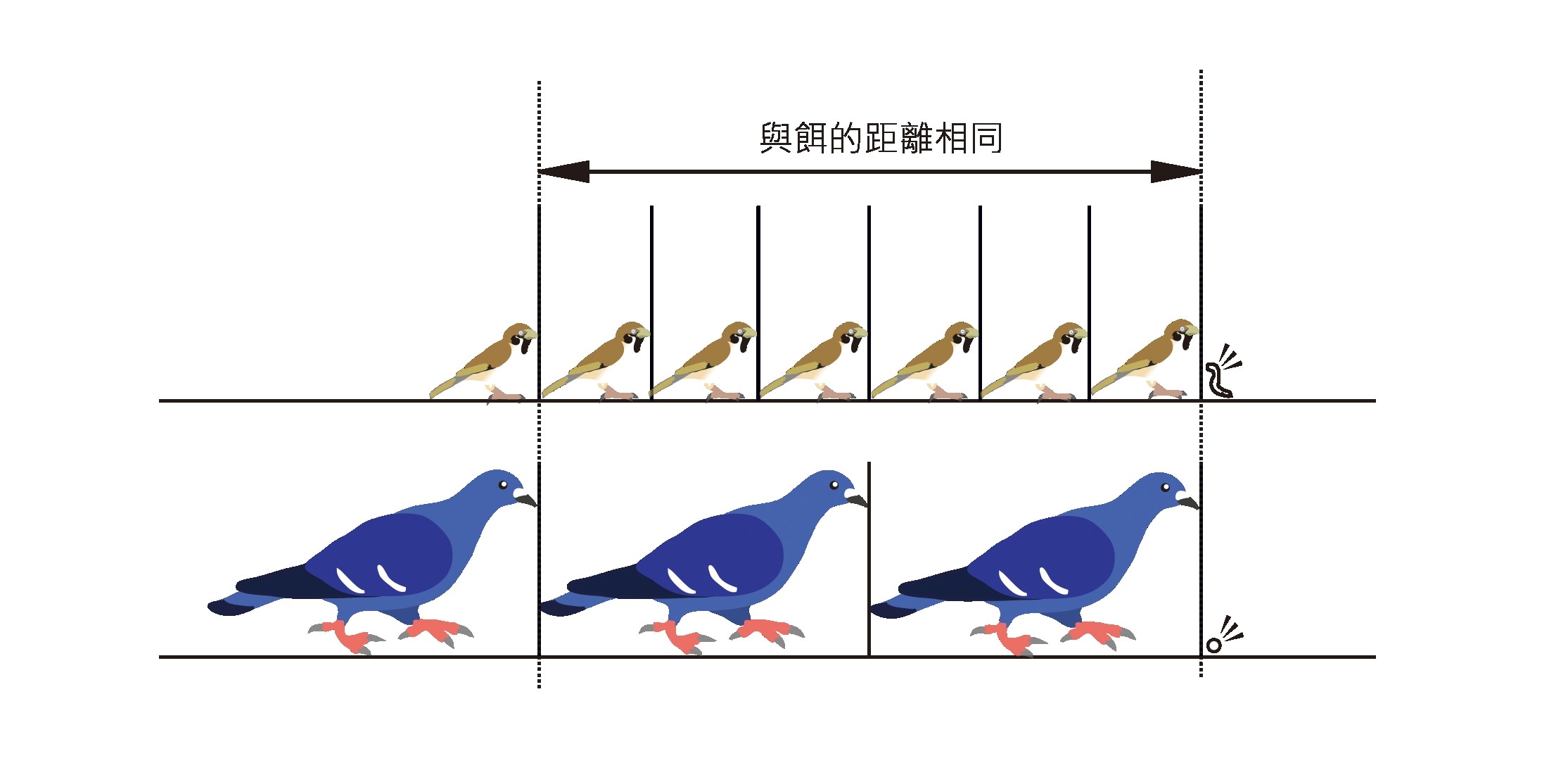
但依筆者經驗,最常見的錯誤防治法就是在面積不小的玻璃上只貼一張猛禽貼紙或鷹眼貼紙,認為鷹的形象可以嚇阻鳥兒不接近,但最後卻發現效果不彰。

其實,這就如同在農田設立稻草人,鳥類會判斷環境中的威脅者,當牠發現貼在玻璃的飛鷹不會動,會判定沒有威脅,自然不當一回事,反而想從沒貼貼紙的玻璃處飛去而導致窗殺。
因此美國奧杜邦學會曾做過研究,想知道到底要布置多密,野鳥才不會飛撞玻璃。實驗結果顯示,寬 5 公分 × 長 10 公分(約 2 英寸 × 4 英寸)的布置間隔可以防止 90% 以上的窗殺,或許讀者們可以參考,做為野鳥窗擊風險玻璃的改善準則。

延伸閱讀
- Bird-Safe Building Guidelines
- 野鳥撞玻璃回報 (Reports on Bird-Glass Collisions)
- Birds Striking Building Windows Final 032014
〈本文選自《科學月刊》2020 年 3 月號〉
在一個資訊不值錢的時代中,試圖緊握那知識餘溫外,也不忘科學事實和自由價值至上的科普雜誌。