只知 DNA 卻不知型態的丹尼索瓦人
古代DNA 研究發展至今,最大的驚奇或許是丹尼索瓦人(Denisovans)。過往的古人類學,主要靠化石與人造物認識古代人類,例如知名的尼安德塔人(Neanderthals),就有成百上千的遺址以及大批化石,讓專家得以掌握他們的型態特徵與生活方式。等到尼安德塔人的基因組被定序成功,我們對尼安德塔人的認識,更是進一步拓展到遺傳的層次。

智人與尼安德塔人之外,至今所有滅絕的古人類都是只有化石,不知 DNA;而唯一例外是丹尼索瓦人。目前已知丹尼索瓦人的居住地點,只有西伯利亞南部的丹尼索瓦洞穴一處,古遺傳學家由此地出土的一位蘿莉手指的骨頭中,取得了高品質的全套丹尼索瓦基因組。
- 延伸閱讀:丹尼索瓦人(上):尼安德塔人的神秘近親
然而,除了此一小到難以辨識型態的指骨以外,我們對丹尼索瓦人型態的了解只有臼齒特別大,然後就沒有其他資訊了。也就是說,絕大部分古人類是缺乏遺傳訊息,只知化石型態;丹尼索瓦人卻是只知 DNA,不知型態。
看起來,在找到更多丹尼索瓦人的化石之前,我們應該無從得知他們的長相?由耶路撒冷希伯來大學的 Liran Carmel 博士率領的戰隊,卻出乎意料地提出無比瘋狂的主意:即使缺少新的化石證據,靠著表觀遺傳學(epigenetics)仍然有辦法無中生有,重建丹尼索瓦人的面貌。

能調控基因表現的DNA甲基化
基因的 DNA 序列就算沒有改變,細胞也能靠著不同的調控方式,影響基因的表現。透過不同化學修飾,不改變 DNA 本身仍能影響基因表現的方法,稱作表觀遺傳學修飾(epigenetic modification);其中一種方法是在 DNA 鹼基上加上甲基,也就是 DNA甲基化(DNA methylation),基因組上被甲基化的區域,通常會降低或失去表現能力。
即使 DNA 序列一模一樣,若是甲基化程度不同,基因表現仍可以產生差異。智人、尼安德塔人、丹尼索瓦人三者的親戚關係很近,遺傳差異很少,彼此間許多型態上的差別,或許並非 DNA 序列差異造成,而是與基因調控有關,DNA甲基化在其中可能就扮演著重要角色。
問題是我們要怎麼才能知道,古代基因組上 DNA甲基化的狀態?DNA 不會變,但是像 DNA甲基化這般的表觀遺傳學修飾卻會時常變化,而且就算是同一個人,不同細胞、組織、生理狀態之下,基因組上 DNA甲基化的狀況也會有差異;因此,如果要研究 DNA甲基化,一定要注意細胞來自什麼狀態。
古代DNA 更絕望的問題是,也許幾萬年後 DNA 序列仍然生還,但是上頭的化學修飾,一定早就死光光,沒有任何辦法可以復原。
死掉幾萬年,還能得知 DNA甲基化圖譜?
幸好,雖然古代DNA 上的化學修飾沒了,不過衍生結構還在。人類基因組是由 ATCG 排列而成,大部分 DNA甲基化的位置,都位於 CG 連續出現時前面的 C,也就是胞嘧啶(cytosine)上頭,由此可以預測基因組上,哪些位置被甲基化的機率較高。
不過要如何分辨,哪些胞嘧啶曾被甲基化?生物死掉以後 DNA 會開始損壞,而最常見的是去胺作用(deamination),胞嘧啶經過去胺作用後,會轉變成尿嘧啶(uracil,簡稱 U),甲基化的胞嘧啶(5-甲基胞嘧啶)卻會變成胸腺嘧啶(thymine,簡稱 T)。賓果!
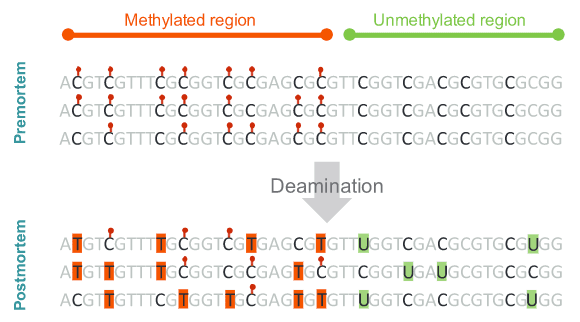
由於化學反應之故,從化石中取得的古代DNA 片段,與智人的基因組對照以後,智人是 C 的位置,古代基因組上若是 U,意謂古人類該位置本來很可能是 C,讀取到 U 是去胺作用所致。假如古代基因組上是 T,那就有兩個可能:第一,它真的是有別於智人的 T;第二,它本來是甲基化的 C,被去胺作用轉變成 T。
用次世代定序法一網打盡樣本內所有 DNA 片段,通通定序的好處是,可以取得許多古代DNA 片段,對應到基因組上的同一個位置;藉此能夠估計古代基因組上,該位置究竟真的是 T,或是甲基化 C 的機率。靠著這套策略,古代基因組上的 DNA甲基化圖譜(DNA methylation map)被順利重建。[1]
探討表觀遺傳學調控很重要的是,不同細胞的修飾狀況不同。古代基因組的 DNA甲基化圖譜,大致看來與智人的骨細胞類似,想來十分合理,因為古代基因組正是取自尼安德塔人,與丹尼索瓦人的骨細胞。
用基因調控差異,重建未知的型態
得知基因組上哪些位置被甲基化以後,接下來是比較智人與尼安德塔人、丹尼索瓦人間的差異,以及探討 DNA甲基化的差異,是否會影響三者的型態特徵。最後,則是重建型態。
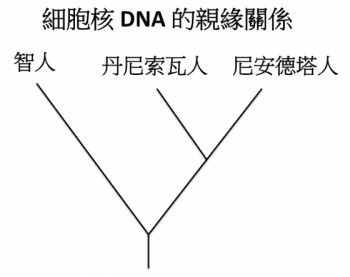
由 DNA 差異判斷,尼安德塔人、丹尼索瓦人的血緣關係較近,共同祖先約能追溯到 40 多萬年前,又與智人大概在 55 到 77 萬年前分家。
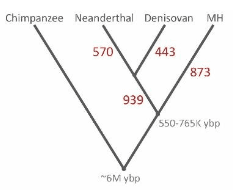
整體看來,基因組上甲基化位置彼此有別的數目,尼安德塔人與丹尼索瓦人分家以後,尼安德塔人有 570 個,丹尼索瓦人則有 443 個。他們的共同祖先與智人分開之後有 939 個,智人這邊則是 873 個。[2]
比對許多資料庫與序列,費了一番功夫後,研究團隊建立起 DNA甲基化、相關基因表現,與臉、聲道、咽、喉型態發育的關係。尼安德塔人有大量化石,型態如何是已知的;用尼安德塔人的型態特徵,可以回推 DNA甲基化和基因表現的改變,對型態的影響。
假如利用古代基因組的 DNA甲基化圖譜,能夠順利重建尼安德塔人的臉部型態,而丹尼索瓦人相關的基因調控狀況,又與尼安德塔人和智人相差不多,那麼同樣的預測辦法,就能用於重建丹尼索瓦人的面貌。結果尼安德塔人型態的預測,正確率約有 87%,可見預測大致上是可靠的。
於是,研究團隊重建了丹尼索瓦人的型態。假如跟想的一樣,丹尼索瓦人會有不少特徵與尼安德塔人一致,不過也有 56 項有別於智人和尼安德塔人,例如丹尼索瓦人配備更長的牙弓(dental arch),以及更寬的雙頂骨(biparietal)。
丹尼索瓦人是「他」?
非常非常有意思的是,研究團隊也將預測的丹尼索瓦人和許昌人比較,而且驚訝地發現,許昌人已知的型態特徵中,大部分都與「丹尼索瓦人」吻合。
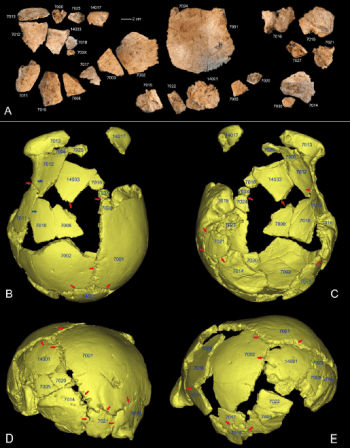
許昌人出土於中國河南省,許昌的靈井遺址,是生活在東亞北方,10 萬年前的古早人類;型態特徵與所有已知古人類都不完全一致,所以難以歸類;還有 2 項特徵與尼安德塔人一致。某些古人類學家認為,既然有共同的型態,許昌人和尼安德塔人間應該有某種糾葛;還有人推論,許昌人多半與丹尼索瓦人有關係,甚至大膽猜測許昌人,就是面貌未知的丹尼索瓦人本尊。[3]
至今為止,用 DNA甲基化圖譜重建古人類型態的研究,都還沒有正式發表。丹尼索瓦人的型態預測是否正確,許昌人是不是就是各界尋覓已久的丹尼索瓦人,這些令人期待的問題,仍有待未來分曉。
至少我們如今知道,聰明的智人已經想到辦法,有望能無中生有,重建滅絕親戚未知的面貌。
延伸閱讀:
- 短篇 臉部形態模擬,顯示智人與尼安德塔人,共同祖先處於70萬年前
- 缺乏遺傳多樣性的孤雌生殖,註定死路一條?——進擊の大理石紋螯蝦(下)
- 我們都有尼安德塔人的血統,但你知道你有多尼安德塔嗎?
- 丹尼索瓦人(下):翻轉人類演化學的古人種
- 不用觀落陰,DNA帶你重回人類大歷史現場 ——古代DNA追追追(上)
參考文獻:
- Gokhman, D., Lavi, E., Prüfer, K., Fraga, M. F., Riancho, J. A., Kelso, J., … & Carmel, L. (2014). Reconstructing the DNA methylation maps of the Neandertal and the Denisovan. Science, 1250368.
- Extensive Regulatory Changes in Genes Affecting Vocal and Facial Anatomy Separate Modern from Archaic Humans
- Li, Z. Y., Wu, X. J., Zhou, L. P., Liu, W., Gao, X., Nian, X. M., & Trinkaus, E. (2017). Late Pleistocene archaic human crania from Xuchang, China. Science, 355(6328), 969-972.
本文亦刊載於作者部落格《盲眼的尼安德塔石匠》暨其 facebook 同名專頁。