

東亞人常見,美洲原住民都有的「箕形門齒」
大家或許有聽過「箕形門齒(shovel-shaped incisors)」,箕是鏟子的意思,簡單來說,就是門齒上有個像是鏟子形的型態。有些資料會說箕形門齒是「漢人」的特徵,嗯……不要太吹毛求疵的話,倒也不是不能這樣講,只是這樣說的意義不大;還有些資料會說是「純種漢人」特色,這就是無稽之談了。
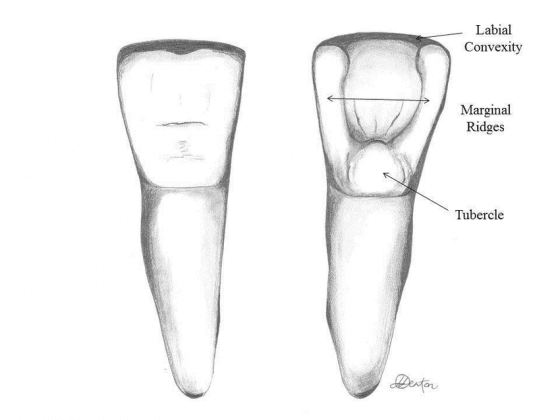
另外也有個流傳已久的傳統論點:東亞的現代智人和古代的直立人(如北京人)都配備箕形門齒,因此兩者間必有血緣關係。這部分我們之後再來討論,先來看比較確定的事實。
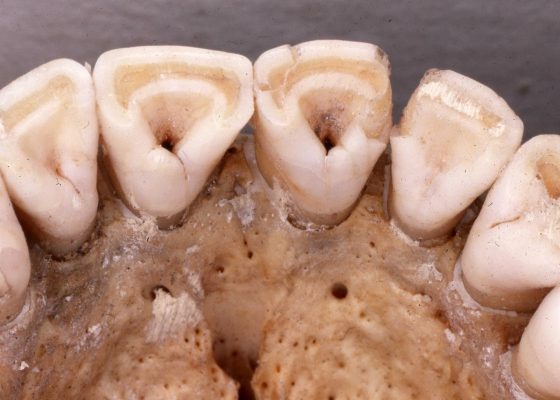
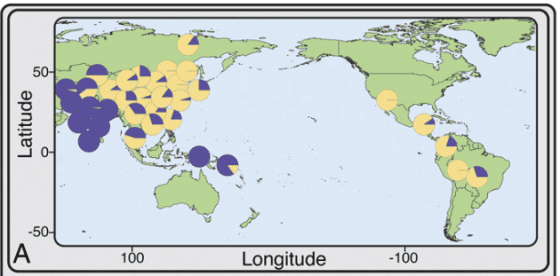
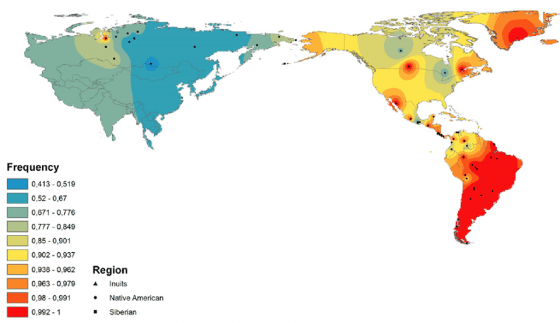
箕形門齒廣泛分佈在現代的東亞族群中,若以現代族群分類歸納的話,箕形門齒並不是「漢人」的專利,韓國、日本人配備此一型態門齒的比例也相當高;整體來看,現代東亞族群約有 40% 的人擁有箕形門齒。不只有現代,此一特徵在東亞大陸各處、或是台灣的史前遺址(如烏山頭遺址 [1])中也很常見;能說是從古到今,很多東亞居民都擁有箕形門齒。
儘管如此廣泛,東亞人擁有箕形門齒的比例還遠不及美洲原住民;在尚未與歐洲人接觸前的美洲族群,有箕形門齒特徵的比例逼近100%。而在非洲與歐洲族群中,箕形門齒的比例則非常非常低;所以此一牙齒型態,算是所有美洲原住民與部分東亞人,有別於其他族群的特色。[2]
箕形門齒與 EDAR 基因之間的糾葛
從上述我們可知,牙齒的型態在不同的族群會有所不同,但究竟箕形門齒是如何產生的?
遺傳學研究指出箕形門齒比較不受環境影響,不過它與EDAR(全名 ectodysplasin A receptor)基因高度相關:假如此基因序列上的第 370 號氨基酸,由纈胺酸(valine)改變為丙胺酸(alanine),那麼就會讓門齒變成箕形。[3]
非洲、歐洲族群則幾乎不存在箕形門齒,EDAR 基因的第 370 號氨基酸也多半是纈胺酸;而在美洲原住民與東亞族群中,此一位置是丙胺酸(可以以 EDAR V370A 表示)的比例很高。過去曾有研究估計過此一遺傳變異大約在 3 萬年前誕生,世界上兩群人間差異如此巨大,許多學者認為是受到天擇影響。
那到底箕形門齒,是否曾替東亞與美洲人帶來什麼好處呢?
等等等等,為什麼在演化上帶來好處的會是箕形門齒呢?不就是顆怪怪的門齒能有什麼功用呢?基因型與表現型的關係十分複雜,許多表現型受到不只一個基因型影響;反過來說,一個基因型往往也會影響很多表現型。在這個案例中,箕形門齒深受 EDAR 基因變異的影響,然而會被 EDAR 影響的特徵,箕形門齒只是區區一種。
EDAR 基因的蛋白質產物,涉及一條很重要的生化反應路徑:NF-ĸB 訊息傳遞路徑。目前已知 EDAR 的 V370A 變異影響非常廣泛,除了門齒箕形度,還有毛髮、臉部型態(至少耳垂與下巴)、汗腺(密度變高)、乳腺(分枝變多),以及門齒以外的一些牙齒型態等等。EDAR 涉及的每一項特徵,都可能是天擇力量作用的目標。[4][5]
不論住在何地、何時、任何環境的美洲原住民,近乎 100% 擁有箕形門齒,由此可以推論,每個人遺傳上都配備 V370A 變異。為什麼比例會如此極端地高呢?合理的假設是,EDAR 基因一度對於美洲原住民的祖先十分重要,只有配備 V370A 變異的個體才能留下血脈,沒有的都在歷史洪流中慘遭淘汰了。
問題是,受到 EDAR 基因型影響的表現型這麼多,哪一項才有那麼大的影響力?
最近發表的論文認為,問題關鍵在:母乳!

當美洲原住民的祖先,仍住在冰河時期的白令時
在談論為什麼是母乳前,讓我們先話說從頭:所有美洲原住民共享的特徵,很可能可以追溯到所有美洲族群尚未分家以前,共同祖先的時期。遺傳學研究估計,美洲原住民與西伯利亞族群大概分家 2 萬多年;那時是冰河時期,白令地區還不是海峽,而是可以直接走路通過的陸橋。[6][7]
在此之後,美洲原住民的祖先由白令進入美洲。有些跡象顯示,美洲原住民的祖先在白令地區住了相當一段時間,或許有好幾千年之久,之後才大舉南遷,四散各地。冰河時期白令的環境,很可能就是關鍵。

陽光中的紫外線與維生素D 有關,而維生素D 對健康不可或缺;一般狀況下,人的皮膚吸收紫外線後,能自行生產足夠的維生素D。人類祖先離開非洲,移民高緯度之後膚色變淡,和高緯度陽光弱,皮膚需要更多紫外線生產維生素D,應該很有關係。
然而,白令的緯度更高、陽光更弱,幾乎沒什麼紫外線;假如長期住在此處,膚色再淡恐怕也無法彌補紫外線過低的問題。所幸即使沒有日曬,人類仍能由飲食中獲取維生素D,美洲原住民的祖先藉由飲食補充維生素D,得以免除滅團的命運。
在沒有陽光的日子,寶寶靠母乳補充維生素D
但是有一種人沒辦法吃東西,那就是剛出生的寶寶。營養匱乏的古代,母乳對寶寶發育相當重要,在白令可能更加重要,因為母乳是寶寶唯一的營養來源;無法吃東西的寶寶,只能透過母乳攝取維生素D 維持健康、順利長大。女生的 EDAR 基因若是配備 V370A 變異,乳腺分枝會變多,將能分泌更多母乳。
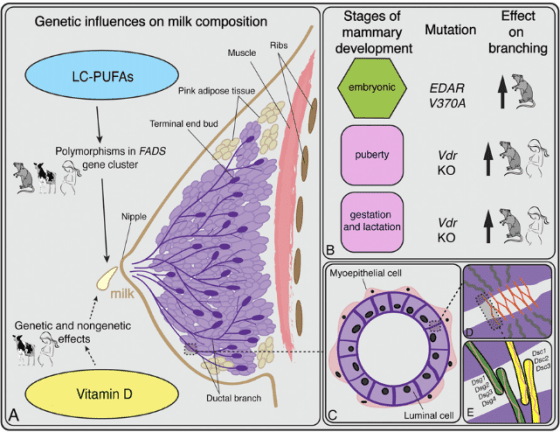
論文推論,在白令特殊的環境下,能增量母乳的遺傳變異 EDAR V370A,將能帶來無與倫比的演化優勢,因此席捲了 2 萬多年前,住在白令的小小族群;而箕形門齒,是配備此變異的間接產物。等到後來白令居民南遷,分家發展出美洲各地的族群以後,即使環境已經與當年不同,仍保持此一特徵。
- 延伸閱讀:懷孕期間補充維生素 D 能反應到母乳中
要提醒各位讀者的是,這套觀點講起來合理,不過仍需更多證據支持。論文提出一項佐證是,還有一個基因與 EDAR 狀況類似,那就是負責合成不飽和脂肪酸的「脂肪酸去飽和酶 FADS」(全名 fatty acid desaturase)。
也許還要脂肪酸?
2014 年發表的論文報告,住在格陵蘭的伊努特人,去飽和酶的基因序列和中國漢人、歐洲人不太一樣。那時推論是,伊努特人此一基因受到天擇作用,有助於他們適應缺乏穀物,富含大量動物性油脂的高油飲食。[8]
然而 2017 年發表的論文,驚覺上述假說有問題!不論住在哪裡,幾乎所有美洲原住民族群中,都有很高比例的去飽和酶基因配備該變異;可是只有伊努特人住在冰天雪地,吃很多魚和海豹,其他美洲居民卻都不是,表示美洲族群的去飽和酶,和其他人的差異,或許與伊努特人的特殊環境無關。[9]
會不會與白令經歷有關呢?去飽和酶基因改變,會影響不飽和脂肪酸的生成。不飽和脂肪酸,如 DHA、EPA,會影響神經發育與許多生理反應,這回論文的推論是,美洲原住民祖先還住在白令時,不飽和脂肪酸也相當關鍵。和 EDAR 一樣,在冰河時期的特殊環境中,有利的去飽和酶遺傳變異,席捲白令的小小族群,使得特定版本的去飽和酶和 EDAR 一樣,成為日後美洲原住民的標準配備。
至於是怎麼個影響法,目前不適合遽下定論。EDAR 一個基因就會影響許多表現特徵,反過來說,就是很多外在表現都有機會影響 EDAR 的演化,而去飽和酶也是如此,根據已知線索,仍難以判斷天擇對去飽和酶的作用目標。
EDAR 基因 x 箕形門齒 x 北京人?
東亞族群中,EDAR V370A 的比例很高,應該也是受天擇影響所致,不過原因未必與美洲原住民相同。如前所述,EDAR 受到許多生理、型態特徵影響,在不同年代、不同環境條件之下,未必只會受到一種外力驅使。
如今可以確定,智人的箕形門齒與 EDAR 基因息息相關。遺傳學估計此一變異起源於數萬年前,若估計正確,那麼智人的箕形門齒,和直立人這類非智人的古人類親戚,之間大概沒什麼直接關係。當然還是有遠古混血的可能性,儘管機率很小,目前仍無法排除。
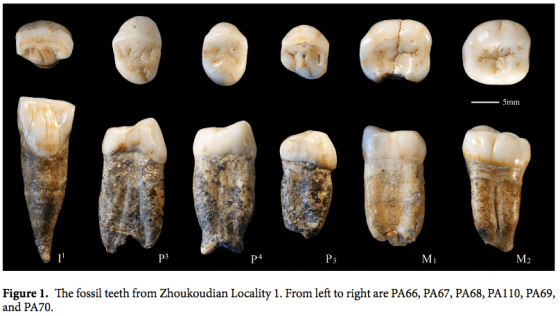
純論型態,今年有一篇全新發表、關於北京人的論文〈The fossil teeth of the Peking Man〉,研究納入了許多非智人與智人族群與北京人一起比較,分析牙齒的型態差異。論文內文有提到北京人化石中,門齒的箕形化(shoveling),也提到此一型態亦見於其他東亞直立人,例如和縣(Hexian)人的化石。[10]
然而,整篇論文完全沒有提及北京人的箕形門齒,與現代東亞族群有任何關聯性。我不是型態專家,不過由這篇論文連一句都不提看來,古代直立人,與今日東亞人的箕形門齒比較,或許並非正經的古人類學家目前關心的問題。
智人的門齒是否箕形受到 EDAR 高度影響,我們不知道直立人是否也是如此,不過智人與直立人差異不是太大,假如直立人的 EDAR 基因影響力和智人類似,也不是太過驚奇(例如對汗腺、毛髮的影響,連用老鼠當實驗動物都能見到)。
由 EDAR 在智人的演化狀況推論,很多因素都有機會間接造成箕形門齒,因此即使觀察到不同族群,都有箕形門齒此一型態,也無法證實它們有著共同的起源,甚至不需要有一樣的起因。
劃重點:
1. 門齒箕形與否,受 EDAR 基因變異影響。
2. 美洲原住民 100% 配備箕形門齒,或許能追溯到冰河時期,東亞版 EDAR 能增加母乳產量所致。
3. 美洲原住民的箕形門齒,是天擇作用後的間接影響。
4. 美洲族群中的脂肪酸去飽和酶,可能和 EDAR 基因狀況類似。
5. 東亞直立人,如北京人的箕形門齒,很可能與智人沒有關係。
6. 人類的演化歷史,何其複雜。
延伸閱讀:
參考文獻:
- 1. 台灣史前文化的奧祕:烏山頭的史前人
- 2. Hlusko, L. J., Carlson, J. P., Chaplin, G., Elias, S. A., Hoffecker, J. F., Huffman, M., … & Scott, G. R. (2018). Environmental selection during the last ice age on the mother-to-infant transmission of vitamin D and fatty acids through breast milk. Proce
- 3. Park, J. H., Yamaguchi, T., Watanabe, C., Kawaguchi, A., Haneji, K., Takeda, M., … & Hanihara, T. (2012). Effects of an Asian-specific nonsynonymous EDAR variant on multiple dental traits. Journal of human genetics, 57(8), 508.
- 4. Kamberov, Y. G., Wang, S., Tan, J., Gerbault, P., Wark, A., Tan, L., … & Powell, A. (2013). Modeling recent human evolution in mice by expression of a selected EDAR variant. Cell, 152(4), 691-702.
- 5. Peng, Q., Li, J., Tan, J., Yang, Y., Zhang, M., Wu, S., … & Jiao, Y. (2016). EDARV370A associated facial characteristics in Uyghur population revealing further pleiotropic effects. Human genetics, 135(1), 99-108.
- 6. Raghavan, M., Steinrücken, M., Harris, K., Schiffels, S., Rasmussen, S., DeGiorgio, M., … & Eriksson, A. (2015). Genomic evidence for the Pleistocene and recent population history of Native Americans. Science, 349(6250), aab3884.
- 7. Moreno-Mayar, J. V., Potter, B. A., Vinner, L., Steinrücken, M., Rasmussen, S., Terhorst, J., … & Reuther, J. D. (2018). Terminal Pleistocene Alaskan genome reveals first founding population of Native Americans. Nature, 553(7687), 203.
- 8. Fumagalli, M., Moltke, I., Grarup, N., Racimo, F., Bjerregaard, P., Jørgensen, M. E., … & Christensen, C. (2015). Greenlandic Inuit show genetic signatures of diet and climate adaptation. Science, 349(6254), 1343-1347.
- 9. Amorim, C. E. G., Nunes, K., Meyer, D., Comas, D., Bortolini, M. C., Salzano, F. M., & Hünemeier, T. (2017). Genetic signature of natural selection in first Americans. Proceedings of the National Academy of Sciences, 114(9), 2195-2199.
- 10. Xing, S., Martinón-Torres, M., & de Castro, J. M. B. (2018). The fossil teeth of the Peking Man. Scientific reports, 8(1), 2066.
本文亦刊載於作者部落格《盲眼的尼安德塔石匠》暨其 facebook 同名專頁。








































