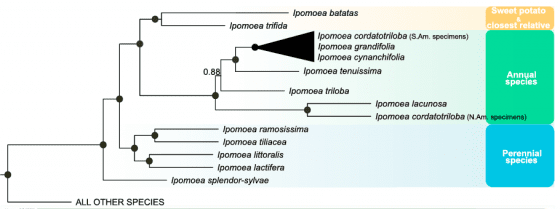
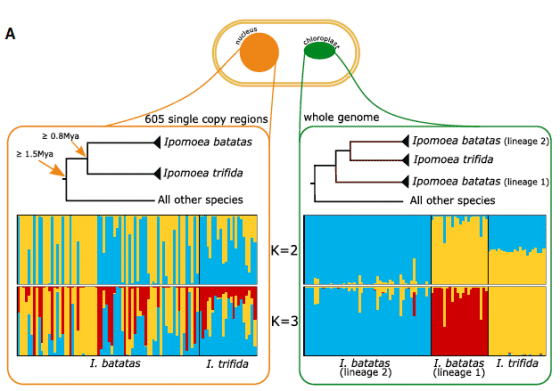
然而,比較親戚間的遺傳差異以後,番薯的六套 DNA 彼此間都更為接近,沒有見到上述狀況。可見番薯的染色體應該只有單一來源,並未經歷過異種合體。家族中所有成員,與番薯最接近的是大星牽牛(Ipomoea trifida, 上上圖B),由此推論,番薯應該是由二倍體的大星牽牛演化而成的「同源多倍體(autopolyploidy)」。
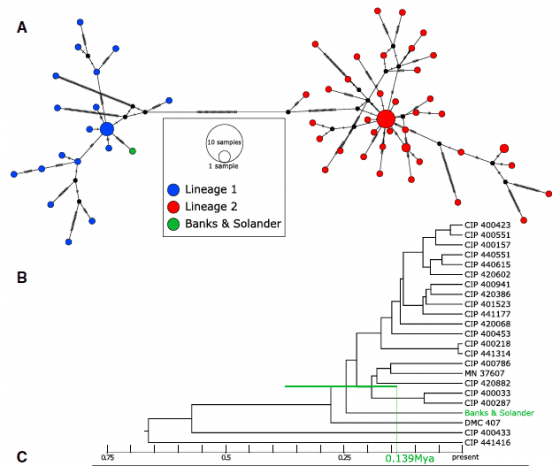
第一部分的質疑點在於,古代番薯樣本是否經過可靠的程序處理?新研究的根據來自古代番薯樣本,這個番薯不是普通的番薯,而是由庫克船長隊伍中的 Joseph Banks 與 Daniel Solander,1769 年採集自社會群島的珍貴標本。定序 DNA 後,得知其葉綠體型號屬於 CLI,估計與美洲親戚至少在 11.5 萬年前分家,遠遠早於人類出現的年代。
1. Mu, P., Carruthers, T., Wood, J. R., Williams, B. R., Weitemier, K., Kronmiller, B., … & Rausher, M. D. (2018). Reconciling Conflicting Phylogenies in the Origin of Sweet Potato and Dispersal to Polynesia. Current Biology, 28(8), 1246-1256.
2. Scotland, R. W., Munoz, P., & Carruthers, T. (2018). Temporal Dynamics of the Origin and Domestication of Sweet Potato and Implications for Dispersal to Polynesia. bioRxiv, 309799.
3. Thomson, V. A., Lebrasseur, O., Austin, J. J., Hunt, T. L., Burney, D. A., Denham, T., … & Linderholm, A. (2014). Using ancient DNA to study the origins and dispersal of ancestral Polynesian chickens across the Pacific. Proceedings of the National Academy of Sciences, 111(13), 4826-4831.
4. Storey, A. A., & Matisoo-Smith, E. A. (2014). No evidence against Polynesian dispersal of chickens to pre-Columbian South America. Proceedings of the National Academy of Sciences of the United States of America, 111(35), E3583.
5. Gonçalves, V. F., Stenderup, J., Rodrigues-Carvalho, C., Silva, H. P., Gonçalves-Dornelas, H., Líryo, A., … & Willerslev, E. (2013). Identification of Polynesian mtDNA haplogroups in remains of Botocudo Amerindians from Brazil. Proceedings of the National Academy of Sciences, 110(16), 6465-6469.
6. Malaspinas, A. S., Lao, O., Schroeder, H., Rasmussen, M., Raghavan, M., Moltke, I., … & Albrechtsen, A. (2014). Two ancient human genomes reveal Polynesian ancestry among the indigenous Botocudos of Brazil. Current Biology, 24(21), R1035-R1037.
7. Moreno-Mayar, J. V., Rasmussen, S., Seguin-Orlando, A., Rasmussen, M., Liang, M., Flåm, S. T., … & Willerslev, E. (2014). Genome-wide ancestry patterns in Rapanui suggest pre-European admixture with Native Americans. Current Biology, 24(21), 2518-2525.
8. Fehren-Schmitz, L., Jarman, C. L., Harkins, K. M., Kayser, M., Popp, B. N., & Skoglund, P. (2017). Genetic Ancestry of Rapanui before and after European Contact. Current Biology, 27(20), 3209-3215.
12. Barbujani, G., Bertorelle, G., & Chikhi, L. (1998). Evidence for Paleolithic and Neolithic gene flow in Europe. American journal of human genetics, 62(2), 488.