


恐龍的生殖生物學
恐龍的生殖生物學一直是古生物學家研究的重點之一,不僅僅是因為大眾對於恐龍如何生小孩充滿了好奇外,也因為化石紀錄很難保存這些相關的行為或是軟組織,所以更顯得神秘。過去,古生物學家僅能仰賴與恐龍親緣關係最近、且還存活著的兩個類群——鱷類與鳥類,來推測恐龍的生殖生物學。
竊蛋龍:近70年的誤會
幸運的是,其中有一類恐龍,僅生活在白堊紀時期的東亞地區與北美洲,牠們頭上有冠、沒有牙齒,而且自從1920年代在蒙古被發現第一件標本後,因為在牠們旁邊也發現了一窩被認為是原角龍(Protoceratops)的蛋窩(圖一),因而被命名為竊蛋龍(Oviraptor)。這個名字一路被用到1994年,直到美國自然史博物館的古生物學家發表了一窩來自蒙古國的竊蛋龍胚胎蛋(圖二;參考文獻1),才發現事實上1920年代發現的蛋窩,其實是屬於竊蛋龍自己的,從此竊蛋龍搖身一變,從竊盜的蛋小偷變成呵護寶貝的母親。然而由於生物分類學上命名優先權的規定,竊蛋龍即使被證明不是小偷,牠的名字還是不能變。


竊蛋龍的生殖生物學,介於鱷類與鳥類之間
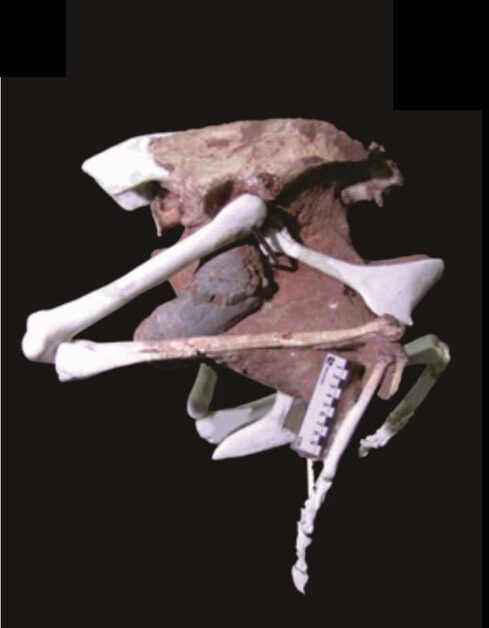
從上述的簡短歷史,各位讀者可能已經發現了:沒有發現帶胚胎的恐龍蛋之前,其實古生物學家是無法得知是什麼恐龍產下這類恐龍蛋的。當然,也有一些例外,例如在2005年,國立自然科學博物館的程延年博士等人,於 Science 期刊上發表了一件內含成對卵的竊蛋龍骨盆(圖三;參考文獻2),在蛋殼上面並沒有看到受酸侵蝕的痕跡,因此程延年博士等人推論,這兩顆蛋應該是還沒被生下來。
而且根據蛋的體積來看,他們也推論竊蛋龍可能跟鱷類相同,有成對的輸卵管(現生鳥類幾乎都只剩下單邊輸卵管);而一邊輸卵管中僅有一顆蛋,這點則與現生鳥類相同。這件珍貴的標本說明:竊蛋龍的生殖生物學是介於鱷類與鳥類之間的過渡型態。
然而,即使有如此漂亮的標本,胚胎蛋的發現,仍然是恐龍生殖生物學最直接的證據。過去也有許多胚胎蛋的發現,例如發現自中國河南省的「路易貝貝」(參考文獻 3)、又或者國立自然科學博物館的楊子睿博士曾在2020年發表的三顆呈現出不同的發育程度的竊蛋龍胚胎蛋(參考文獻 4)、以及一隻竊蛋龍成體趴伏於整窩胚胎蛋上(參考文獻5),說明了牠們不同步孵化的生殖行為策略,早在鳥類之前便開始使用了。
一件發現自江西的絕美胚胎
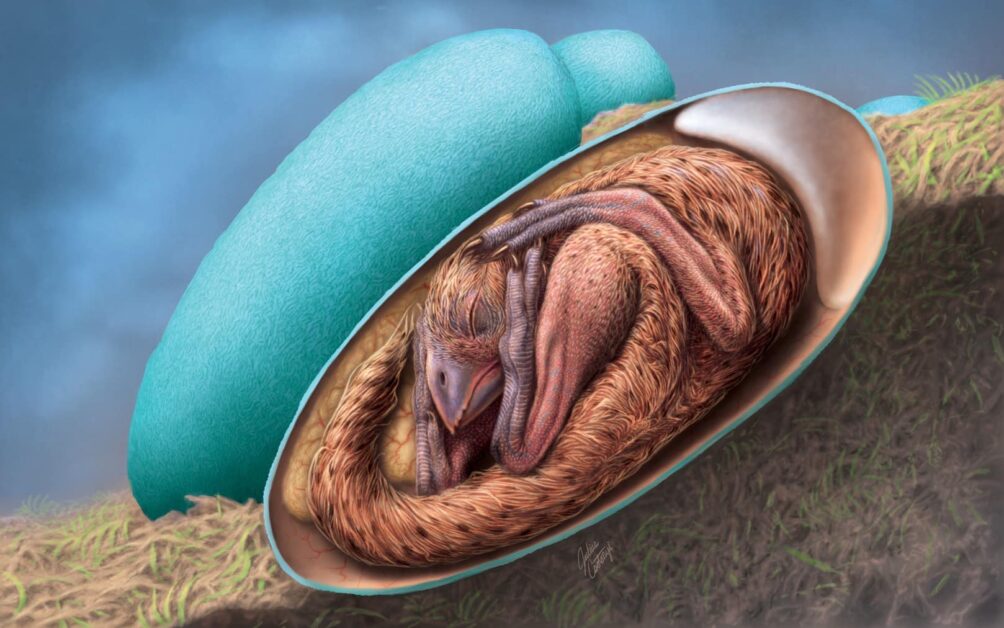
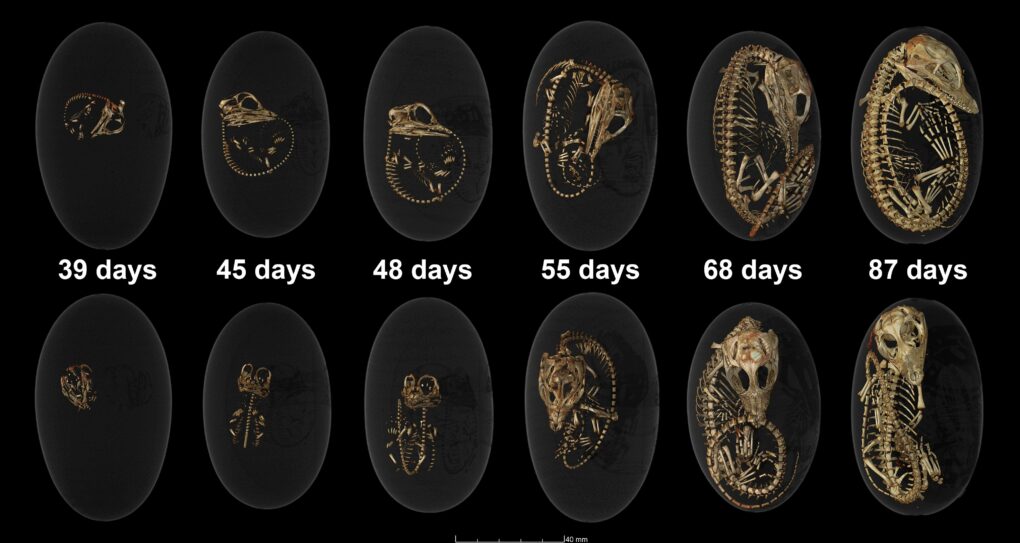
12月21日在 iScience 期刊上,國立自然科學博物館的楊子睿博士與多國的古生物學家合作,報導了一件發現自江西贛州,絕美保存的竊蛋龍胚胎(圖四;參考文獻4)該件標本是老鄉於江西贛州地區發現以後,輾轉到了中國福建省的英良石材自然歷史博物館,經過精細的清修才顯現出完美的胚胎骨骼。這件胚胎的保存狀態,遠比以前曾經報導過的所有竊蛋龍胚胎來得完整,且其發育階段更接近孵化的狀態。

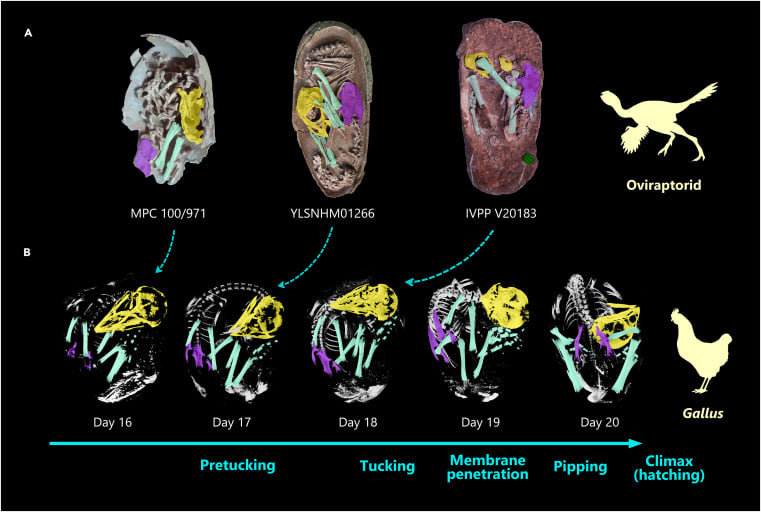
根據骨骼的接合程度(articulation),我們認為這件標本的狀態大約是發育了75%。最特別的是,隱隱約約地表現出將頭顱擺放在右前肢內(這樣的動作稱為 tucking,見圖五復原圖),有如鳥類睡覺姿態一般。這樣的胚胎姿態在主龍類中(Archosauria,包含翼龍、鱷類、恐龍以及鳥類),之前只有現生鳥類胚胎與中生代的反鳥類胚胎有相關的證據。而恐龍現生的另外一支親戚——鱷類,其胚胎在出生前僅會垂至胸前,與鳥類大不相同(見圖六),其中一個最大的原因就來自於鳥類可收可折的前肢結構,所以也不意外鳥類的祖先們——恐龍的胚胎會有類似的情況。
恐龍比鳥類更早知道:孵化前把頭埋進腋窩,順產率更高
鳥類學家研究發現,鳥胚胎在出生前若無法順利的將頭顱摺入右前肢當中,便有很高的機率孵化失敗,胎死「蛋」中。因此,將頭顱摺入右前肢當中的這個行為,是鳥類胚胎成功孵化的重要因素之一,同時也可能就是牠們能夠熬過白堊紀末的大滅絕事件且存活下來的其中一個優勢。
在Norell 等人1994年的首次報導竊蛋龍胚胎文章中,他們曾經提到過他認為他研究的胚胎姿勢比較像鱷類(圖七)。本次筆者參與的這篇文章便提出不同於 1994 的論點,我們認為其實竊蛋龍也不是那麼像鱷類,因為這件竊蛋龍胚胎的頭看起來幾乎都快要埋進去了。
然而,這同時也是這篇文章的一點缺憾,因為這個胚胎其實也沒有到真正「最後最後」的階段,所以我們很難證明這個胚胎是處在「準備完全把頭塞進去,跟鳥類胚胎一樣」或是「其實就是跟鱷類胚胎差不多」的情形。 不過如何,這樣的標本幫助我們更進一步了解竊蛋龍的胚胎生物學與發育生物學,是很有趣的發現,也期待未來有更多的恐龍胚胎的發現!
參考文獻
- Norell, M. A., Clark, J. M., Demberelyin, D., Rinchen, B., Chiappe, L. M., Davidson, A. R., McKenna, M. C., Altangerel, P., Novacek, M. J. 1994. A theropod dinosaur embryo and the affinities of the flaming cliffs dinosaur eggs. Science 266:779-782.
- Sato, T., Cheng, Y.-N., Wu, X., Zelenitsky, D. K., Hsiao, Y.-F. 2005. A pair of shelled eggs inside a female dinosaur. Science 308: 375.
- Pu, H., Zelenitsky, D. K., Lü, J., Currie, P, J., Carpenter, K., Xu, L., Koppelhus, E. B., Jia, S., Xiao, L., Chuang, H., Li, T., Kundrát, M., Shen, C. 2017. Perinate and eggs of a giant caenagnathid dinosaur from the Late Cretaceous of central China. Nature Communication 8:14952.
- Yang, T.-R., Engler, T., Lallensack, J. N., Samathi, A. Makowska, M., Schillinger, B. 2021. Hatching asynchrony in oviraptorid dinosaurs sheds light on their unique nesting biology. Integrative Organismal Biology 1:obz030.
- Bi, S., Amiot, R., de Fabrègues, C. P., Pittman, N., Lamanna, M. C., Yu, Y, Yu, C., Yang, T.-R., Zhang, S., Zhao, Q., Xu, X. 2021. An oviraptorid preserved atop an embryo-bearing egg clutch sheds light on the reproductive biology of non-avialan theropod dinosaurs. Science Bulletin 66:947-954.
- Xing, L., Niu, K., Ma, W., Zelenitsky, D. K., Yang, T.-R., Brusatte, S. L. 2021. An exquisitely preserved in-ovo theropod dinosaur embryo sheds light on avian-like prehatching postures. iScience (in press)








































{kind=link}