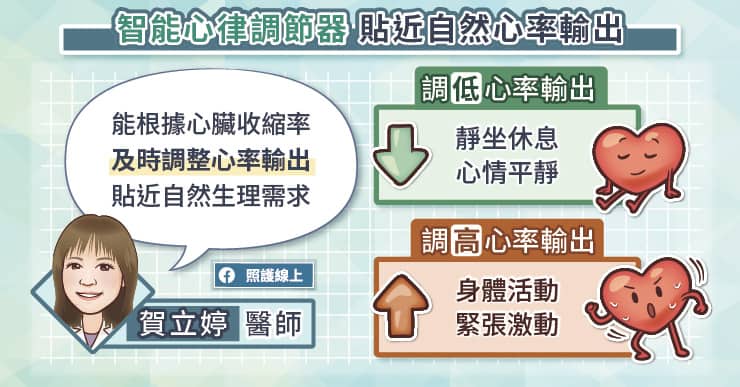
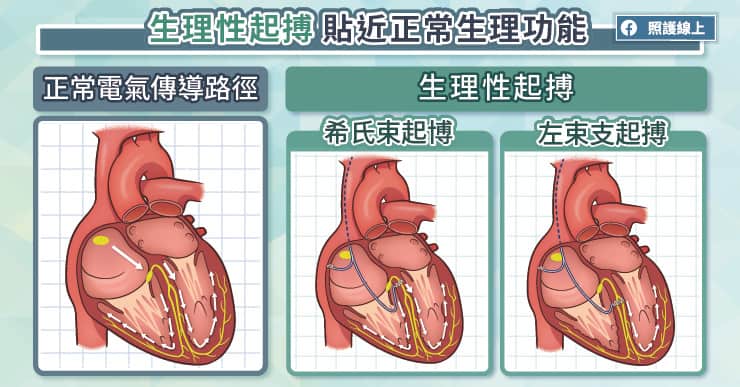
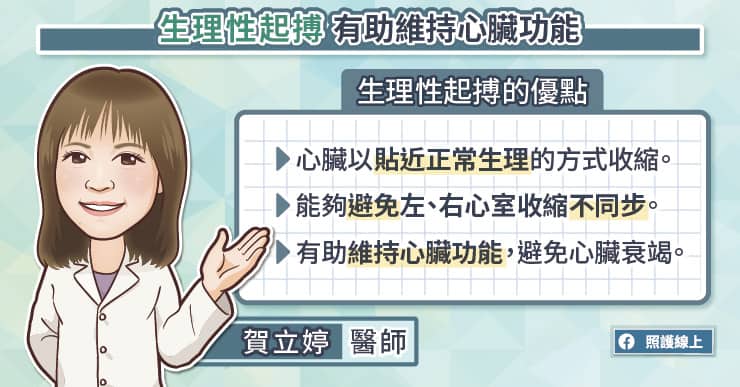
隨心所至並不容易,但嬰兒和媽媽卻只要互相投以微笑,就能跟隨對方的心跳。一項新的研究指出,三個月大的嬰兒,和他的母親能彼此同步心跳,誤差在僅僅幾毫秒內。科學家找來40對母子,利用皮膚電極貼片,追蹤心跳。發現只要一個愛的眼神相會,或者柔情的低語,心跳幾乎能立刻同步。藉由怡人的互動同步心跳,目前只發現存在於媽媽和嬰兒間(譯註:研究包含三個月到13歲大的孩童),研究成果發表於《嬰兒行為及發展》(Infant Behavior and Development)期刊。科學家推測,當人類接收到來自另一個體的表情語言,會開啟腦中特定區域,並調整心跳研究團隊過去的研究也顯示,無法接收來自母親聲音的嬰兒,會像青少年一樣較無法產生情感上的共鳴。早熟的嬰兒,或有產後憂鬱的母親,有高風險會喪失社交技巧,因為他們可能錯過早期母子互動的機會。
隨心所至並不容易,但嬰兒和媽媽卻只要互相投以微笑,就能跟隨對方的心跳。一項新的研究指出,三個月大的嬰兒,和他的母親能彼此同步心跳,誤差在僅僅幾毫秒內。科學家找來40對母子,利用皮膚電極貼片,追蹤心跳。發現只要一個愛的眼神相會,或者柔情的低語,心跳幾乎能立刻同步。藉由怡人的互動同步心跳,目前只發現存在於媽媽和嬰兒間(譯註:研究包含三個月到13歲大的孩童),研究成果發表於《嬰兒行為及發展》(Infant Behavior and Development)期刊。科學家推測,當人類接收到來自另一個體的表情語言,會開啟腦中特定區域,並調整心跳研究團隊過去的研究也顯示,無法接收來自母親聲音的嬰兒,會像青少年一樣較無法產生情感上的共鳴。早熟的嬰兒,或有產後憂鬱的母親,有高風險會喪失社交技巧,因為他們可能錯過早期母子互動的機會。


 隨心所至並不容易,但嬰兒和媽媽卻只要互相投以微笑,就能跟隨對方的心跳。一項新的研究指出,三個月大的嬰兒,和他的母親能彼此同步心跳,誤差在僅僅幾毫秒內。科學家找來40對母子,利用皮膚電極貼片,追蹤心跳。發現只要一個愛的眼神相會,或者柔情的低語,心跳幾乎能立刻同步。藉由怡人的互動同步心跳,目前只發現存在於媽媽和嬰兒間(譯註:研究包含三個月到13歲大的孩童),研究成果發表於《嬰兒行為及發展》(Infant Behavior and Development)期刊。科學家推測,當人類接收到來自另一個體的表情語言,會開啟腦中特定區域,並調整心跳研究團隊過去的研究也顯示,無法接收來自母親聲音的嬰兒,會像青少年一樣較無法產生情感上的共鳴。早熟的嬰兒,或有產後憂鬱的母親,有高風險會喪失社交技巧,因為他們可能錯過早期母子互動的機會。
隨心所至並不容易,但嬰兒和媽媽卻只要互相投以微笑,就能跟隨對方的心跳。一項新的研究指出,三個月大的嬰兒,和他的母親能彼此同步心跳,誤差在僅僅幾毫秒內。科學家找來40對母子,利用皮膚電極貼片,追蹤心跳。發現只要一個愛的眼神相會,或者柔情的低語,心跳幾乎能立刻同步。藉由怡人的互動同步心跳,目前只發現存在於媽媽和嬰兒間(譯註:研究包含三個月到13歲大的孩童),研究成果發表於《嬰兒行為及發展》(Infant Behavior and Development)期刊。科學家推測,當人類接收到來自另一個體的表情語言,會開啟腦中特定區域,並調整心跳研究團隊過去的研究也顯示,無法接收來自母親聲音的嬰兒,會像青少年一樣較無法產生情感上的共鳴。早熟的嬰兒,或有產後憂鬱的母親,有高風險會喪失社交技巧,因為他們可能錯過早期母子互動的機會。