目前全球近7000種的兩棲類動物中,約40%的物種面臨生存上的威脅,正在逐漸地消逝中。自1970年代以來科學家開始發現各大洲(當然沒有南極洲)的蛙類物種都出現了大量暴斃的現象,但死因終究不明。直到1999年學界才發現到這樣種喪心病狂的殺戮高手,是一種命名為蛙壺菌(Batrachochytrium dendrobatidis,以下簡稱Bd)的真菌。染上蛙壺菌的青蛙皮膚會開始大量增生角蛋白引發皮膚病變,影響離子、滲透壓、呼吸等正常生理機能,最後死去,致死率高達8成。更可怕的是,這種疾病的傳染性高、散播速度也快,幾乎橫掃了全世界熱帶、亞熱帶的兩棲類物種 [註1],2009年Kriger和Hero便指出當時已有287種兩棲類會感染Bd。我們也普遍認同這40年來兩棲類多樣性的驟降與蛙壺菌脫不了關係。

Photograph by Joe Sartore
聽起來慘不忍睹對不對?就在大家傷透腦筋、想盡辦法降低蛙壺菌病的盛行率時,2008年開始,荷蘭的棒紋真螈(Salamandra salamandra terrestris)開始一隻隻地離奇死亡,原先穩定健康的族群到了2011年竟然只剩下4% 個體!因此比利時和荷蘭的研究團隊開始抽絲剝繭,想釐清這樁奇案是否又是蛙壺菌搞的鬼。檢測結果排除了萬惡的蛙壺菌以及在英國肆虐青蛙的蛙虹彩病毒(Ranavirus),命案現場的水文、土壤、物候等環境因子也沒有異常狀況。

Photograph by Richardson Winch
2013年八月發表於PNAS的文章,An Martel等人分離出病原株並且透過系統發生檢測,這樁感染真螈的兇手是個生面孔,一種與蛙壺菌互為姊妹種的蠑螈壺菌(阿就都給你們玩就是了…),學名是Batrachochytrium salamandrivorans 以下簡稱Bs。蠑螈遭受感染後,會再侵蝕皮膚表面,造成組織潰瘍和壞死的病徵,圈養環境下的蠑螈在接觸到Bs後七天內就身亡。咦?那這樣高致病力、高傳染率的Bs不就跟Bd競爭宿主搶得你死我活嗎?不會耶!演化的奧妙之處總是讓你跌破眼鏡。科學家發現Bs與Bd各占鰲頭:Bd專攻熱帶、亞熱帶地區,最適生長溫度是17~25°C之間;Bs主打溫帶攻勢,最適生長溫度是10~15°C之間,5°C也可以長,但在超過25°C的環境下五天內就會敗逃死去。這樣精心設計的棲位分化,一方面讓人讚嘆、另一方面又為我們的兩棲類蒙上一層揮之不去的陰影。
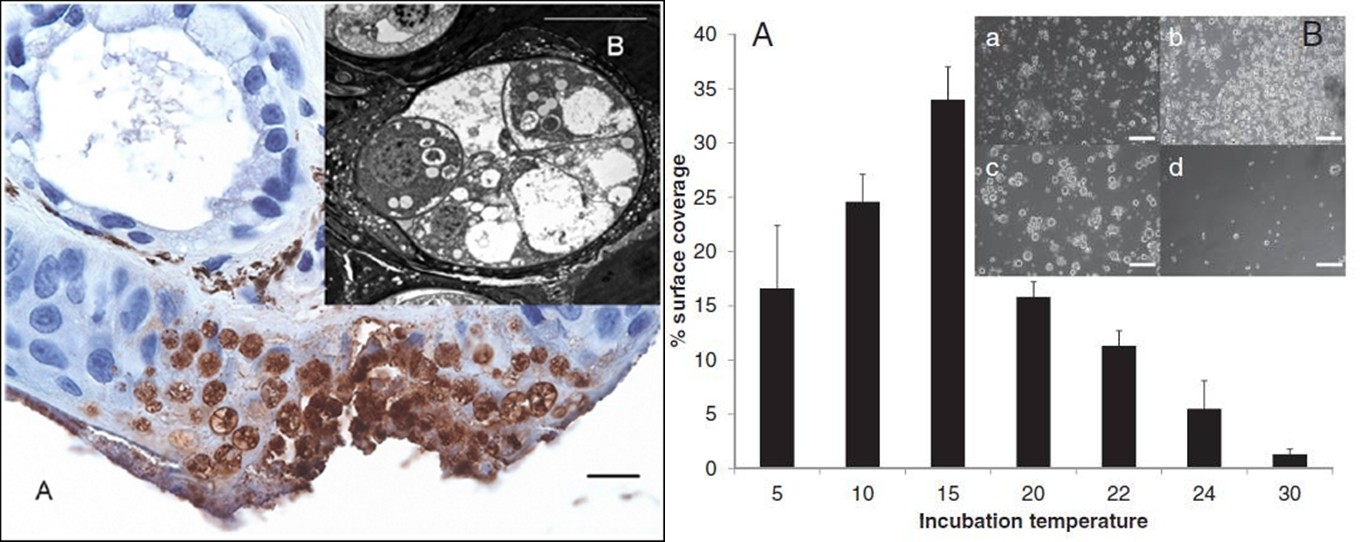
Source: Martel et al. 2009 PNAS
當年這項研究也試圖人工感染當地另一種常見的兩棲類,雄蛙以腳攜卵的產婆蟾(Alytes obstetricans),結果發現Bs沒法感染這種蛙類,團隊因而猜想也許Bs與它的Bd老兄能夠感染蚓螈目、有尾目、無尾目有別,無法感染有尾目外的兩棲動物。
同一個研究團隊(只是作者群從11人變27人了~)在去年(2014) 10月底又發表一篇文章於Science期刊上。為了確認Bs是否真的只會感染有尾目類群,作者找來35種兩棲類動物(蚓螈目1種、有尾目24種、無尾目10種)分別暴露在5000顆Bs孢子下24小時並追蹤個體4個禮拜檢查是否出現病徵,每個禮拜以棉花棒抹取皮膚組織進行qPCR(定量聚合酶連鎖反應)查驗感染程度,感染個體死後再進行組織病理切片檢視患壺菌病情形。這部分的實驗發現只有「有尾目」的成員會受到Bs感染,駭人的是44隻古北區西側的蠑螈中有41隻在染上Bs後迅速地發病死亡。
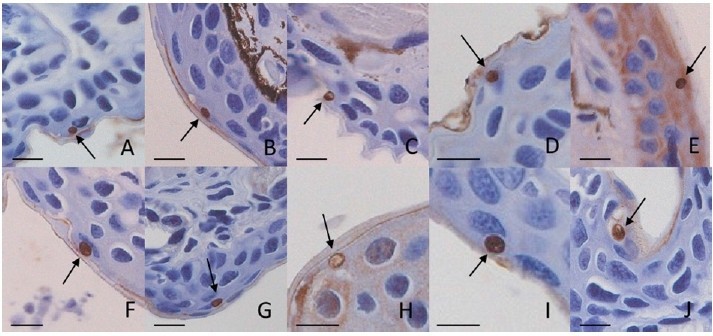
Source: Martel et al. 2014 Science _SM
為了瞭解目前Bs於全球的感染狀況,作者們超大手筆找來了5391隻來自四塊大陸的兩棲動物的皮膚組織,利用qPCR(哦天阿好貴!)掃描這5000多隻樣本有無感染Bs。結果發現東亞(日本、泰國、越南)來的動物皮膚組織上有著和Bs序列一模一樣的DNA,但這些地方不曾有聞為此疾病侵擾。另外在壺菌爆發的疫區,比利時和荷蘭的樣本都有檢驗到Bs的DNA。上述結果意味著蠑螈壺菌可能在亞洲定居已久,當地物種已產生抗性抵禦壺菌,反之,歐洲的蠑螈卻被這些可能近年才引入的真菌搞得人仰馬翻。
接著若依第一階段的感染實驗可將這35個物種受感染表現的不同反應區分為四類,分別為Resistant(不感染不發病)、Tolerant(會感染但沒病)、Susceptible(會感染會發病但是有機會康復)、Lethal(所有受試個體感染後皆發病身亡)。多數歐洲產物種皆為L、多數亞洲產物種為R [註2]、北美產物種則各有R和L。其中最值得一提的是,有三個亞洲產物種的感染反應歸類在S,因此極有可能是Bs的保毒物種,分別是分布日本全境的赤腹蠑螈(Cynops pyrrhogaster)、分布於貴州雲南的藍尾蠑螈(Cynops cyanurus)、分布在越南北部的德氏瘰螈(Paramesotriton deloustali)。本篇從這張系統發生樹圖對應上時間,發現這三種保毒物種共同祖先出現的時間(約4000萬年前,始新世)正好落在Bs與Bd分家時的時間後(約6730萬年前,白惡紀晚期)。

Photograph: Frank Pasmans/PA
由上述結果,Martel等人推斷蠑螈壺菌應是從亞洲起源 [註3],考量現今Bs相當間斷的分布,很有可能是在近幾年隨著人類活動而將Bs帶至歐洲。特別是寵物市場的國際化貿易,每年皆有大量的亞洲產蠑螈被銷往歐美,例如從2001~2009年就有230萬隻東方火龍輸入美國。為此,作者群們又大費周章從歐洲各寵物店、倫敦希斯洛機場、某個香港出口商找來1765隻圈養兩棲類,和570隻其他圈養來源的蠑螈皮膚樣本。檢測發現裡頭有三隻蠑螈為Bs陽性個體,皆為越南疣螈(Tylototriton vietnamensis),追蹤後還發現其中兩隻是在2010被運進歐洲!
究竟這三隻個體是否是把Bs帶入歐洲的元兇,我們需要更多的論證,但可以確定的是全球化的寵物交易與疾病的散播是密不可分的。例如蠑螈壺菌的疫區─荷蘭,除了是出口大量的毒品外,非法寵物貿易也相當盛行,每年都會從南美洲非法進口大量的箭毒蛙。至今,我們似乎還沒有一套有效遏止壺菌病蔓延的方法,在這樣的思維下,各貿易國更應該謹慎地把關世界各地流通的生物,制定輸入與輸出線上的管理、品管等機制,要求各國境內不得將外來種的寵物放生至野外影響當地的生態系統。我們都知道蛙壺菌是怎麼樣撲滅全球的蛙類,如今我們同時得把視角再往溫帶移動,想辦法對付這個新興起的蠑螈壺菌阿。

Photograph by Phùng Mỹ Trung
[註1] 婆羅洲(2011)、巴布亞紐幾內亞(2012) 截至括號內年代尚未有任何感染蛙壺菌的病例。
[註2] 歐洲產但不為L的例外有:掌狀滑螈 Lissotriton helveticus為R。亞洲產但不為R的例外有:三種保毒物種S、西伯利亞極北鯢 Salamandrella keyserlingii為T、文縣疣螈 Tylototriton wenxianensis為L(雖與上述越南疣螈同一屬,但卻歸類在Lethal的等級,也許Bs感染強度也與宿主所棲居的環境因子有關)。
[註3] 2004年Weldon等人指出蛙壺菌Bd可能從非洲起源,他們發現最早在1938年的某隻爪蟾中就檢測到了Bd真菌株。但仍有其他研究認為北美洲東部(Garner et al., 2006)和日本(Goka et al., 2009)也檢測到年代久遠的真菌株,亦有可能是發源地。
An Martel 兩篇文獻:
- An Martel et al. (2013) “Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians” Proceedings of the National Academy of Sciences. 110(38). doi:10.1073/pnas.1307356110
- An Martel et al. (2014) “Recent introduction of a chytrid fungus endangers Western Palearctic salamanders” Science. 346(6209):630-631. doi:10.1126/science.1258268
其他參考文獻:
- Kriger K. M., Hero J. M. (2009) “Chytridiomycosis, Amphibian Extinctions, and Lessons for the Prevention of Future Panzootics” EcoHealth (6)1, 6-10.
- Weldon Ch`e et al. (2004) “Origin of the amphibian chytrid fungus.” Emerging Infect. Dis. 10 (12): 2100–5.
- Kristine K., Grafe T. U. (2011) “Chytrid Fungus Not Found in Preliminary Survey of Lowland Amphibian Populations Across Northwestern Borneo” Herpetological Review 42(1).
- Chris D. et al. (2012) “Batrachochytrium dendrobatidis not found in rainforest frogs along an altitudinal gradient of Papua New Guinea” The Herpetological Journal. 22(3):183-186
- Garner T. W. J. et al. (2006) “The emerging pathogen Batrachochytrium dendrobatidis globally infects introduced populations of the North American bullfrog, Rana catesbeiana.” Biology Letters 2:455–459
- Goka K. et al. (2009) “Amphibian chytridiomycosis in Japan: distribution, haplotypes and possible route of entry into Japan” Molecular Biology. 18(23):4757-4774