

- 作者/張之傑
七月二十一日,清晨拉開窗簾,窗外仍是一座座島嶼,亦即仍在大堡礁範圍之內。
八時五十分左右,和平號停靠凱恩斯港。我們登上十樓甲板繞了一圈,緊靠碼頭處有停車場和公寓,放眼望去,看不到多少高樓,這不是座大城市。碼頭對岸,遠處是起伏的山巒,近處是長堤狀的低矮珊瑚礁,山巒和珊瑚礁之間,隱約可以看出有段水域。
我們參加「前往世界遺產大堡礁綠島航行」行程,十時正集合,隨即下船,步行前往渡輪碼頭,搭渡輪前往綠島。沿著岸邊的棧道前行,十時二十七分到達渡輪碼頭。等候渡輪時,澳洲觀光局的人在發禮物兌換券,可以憑券到市區的禮品店兌換無尾熊玩偶。

綠島是凱恩斯的一座珊瑚島,面積十五公頃,距離凱恩斯約二十七公里,地處世界遺產大堡礁海洋公園範圍內,是個熱門的旅遊景點。該島約形成於六千年前,是波浪將沙子、珊瑚礁等碎屑淤積到珊瑚地基上形成的。
有道是滄海桑田,經年累月的雨水沖洗,沙土中的鹽份逐漸退去,漸漸適合植物生長。風力和鳥類帶來植物的種子,如今島上已有一百二十六種植物。島上沒有淡水,植被仰賴雨水,凱恩斯一帶的雨水足夠支應島上的植物生長。
自二十世紀初以來,旅遊一直是綠島的主要活動。當前的綠島度假村一九九四年開業,有四十六個房間,設有海水淡化廠,二○○一年投入使用,每天可提供五萬五千公升淡水。度假村的游泳池,就是淡水的。
我們等候了約半小時,渡輪才開過來。綠島可能是大堡礁中最容易遊賞的一座島嶼。我們排隊登上渡輪,每人發一套浮潛工具。十一時五十分到達綠島的棧橋碼頭,從開船起到達目的地,航程約四十五分鐘。

棧橋是用一根根木樁撐起來的,長度目測至少一百公尺。下了渡輪,踏上棧道,或許為了防風、防雨或防曬,棧橋的頂部,部份設有半遮式陽棚。我們先到另一艘停舶船(大貓號)上午餐,飯後的半潛艇和玻璃底船活動,都從大貓號上登船。
在大貓號上吃過午餐,十三點十分左右登上半潛艇,透過座位一旁的玻璃窗,觀賞大堡礁海域的珊瑚礁及魚類等動物。解說是事先錄製好的,我們這批乘客以華人為主,所以採用華語錄音。這種解說只是泛泛地談談,不能根據玻璃窗外出現的東西即時說明,所以並沒多大用處。
解說錄音說,珊瑚礁上有很多種海參,具有維護珊瑚礁清潔的作用,海參曾經是澳洲的重要出口海產,綠島當年就是熏製場所。海星中的棘冠海星,觸手六至八隻,身上遍佈棘刺,以珊瑚蟲為食。一九九○年代曾經數量暴增,綠島附近的珊瑚多遭其毒手。我們看到的是正在恢復中的珊瑚礁……。解說錄音沒提白化問題。

我們在半潛艇上約二十五分鐘,回到大貓號,再搭乘玻璃底船出航。半潛艇和玻璃底船的差別,只是前者向玻璃窗外看,後者是扶著圍欄朝腳底下看。至於解說錄音,兩者是同一套,只是玻璃底船的解說沒提到攝影時要將鏡頭貼近玻璃窗。
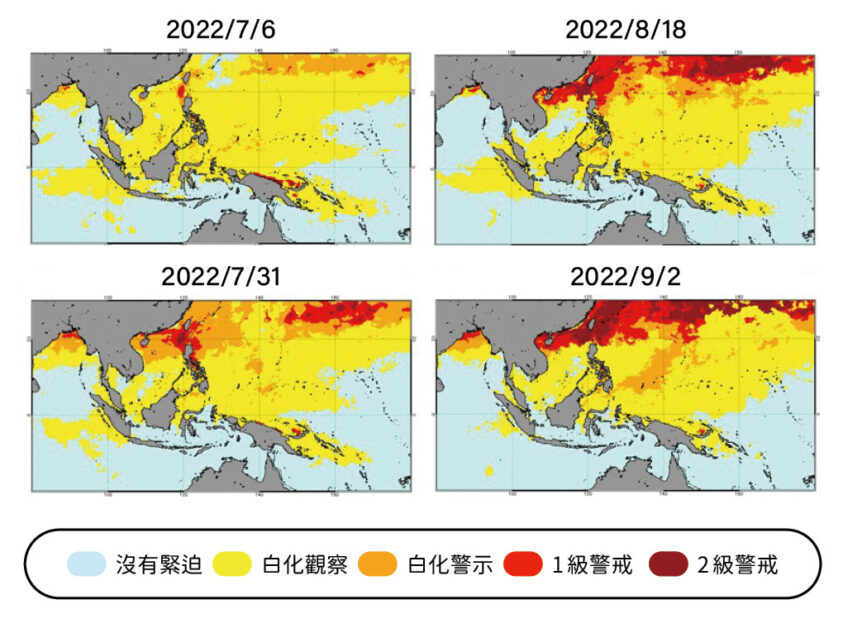
我們在半潛艇和玻璃底船到底看到了什麼?由於污染和氣候變遷等因素,大堡礁的珊瑚約有六成出現白化現象。也就是因為失去共生藻(蟲黃藻),變得灰灰土土,不再色彩絢爛,綠島一帶應該白化得更為嚴重。

從凱恩斯到綠島的渡輪、半潛艇、玻璃底船來來去去,引擎所排出的廢氣哪能不影響到水質!再說,美麗的海葵、帚蟲(屬環節動物)、海蛞蝓等受到騷擾,哪會不縮起來或躲起來!
從前海域沒有污染,在台灣北海岸就能看到絢爛的珊瑚礁世界。大一的普通生物課到基隆附近的八斗子採集,助教不准大夥下海,只能在岸邊的潮池裡活動。隔了一兩個星期,我就自己去了,戴上潛水鏡,眼前出現的景象,簡直就是座海底大花園。這樣的海底大花園想不到連大堡礁都看不到了!
半潛艇和玻璃底船會餵魚,所以魚類倒是看到不少,我對魚類學所知有限,認得出的有成群的鰺魚,和在灰暗的珊瑚礁上繞來繞去的黑帶蝶魚。海龜只看到一隻,好像是蠵龜,是在玻璃底船上看到的。看到海龜,大概是搭乘半潛艇和玻璃底船觀光唯一的收穫。

完成玻璃底船活動,回到大貓號,絕大多數乘客將浮潛工具還給船上。隨即踏上棧道,前往綠島觀光。今天氣溫二十度以下,白色沙灘上仍有些人在作日光浴,還有些人在弄潮戲水。近岸處,從棧橋上向下望,水質清澈見底,珊瑚礁隱約可見。
棧橋入口處,岸邊築有堤防。島上鬱鬱蔥蔥,綠樹掩映下,出現一座木構建築,走近才知道是度假村的紀念品店。度假村紀念品前的小廣場上鋪著木板,顯得特別潔淨,有些樹木已粗可合抱。
資料上說,綠島有大片熱帶雨林。典型的雨林,樹冠分成兩三層,鬱鬱蒼蒼,遮住地面的陽光,地上沒什麼草木。綠島的植被的確有點雨林的樣子。濱海的步道下,有些地方有較大片的沙灘,有些地方緊靠著海。
走到一處緊挨著步道的海灘,我走下步道,想撿點貝殼,卻看到不少浮石。當岩漿噴出地表,壓力驟減,岩漿中的空氣迅速膨脹,熔岩形成多孔性的碎石,氣孔佔體積百分之七十以上。我曾在火山島聖托里尼撿到過浮石,綠島一帶有火山島嗎?
我們十七時三十分回到和平號,上完廁所,洗把臉,稍微整理一下,十七時四十分下船,到市區看看。我們到禮品店換了無尾熊玩偶,又賣了些東西。十八時四十五分回到和平號,放好東西,到四樓餐廳吃定食。回顧這一天,澳洲地大物博,的確是個好地方。不過大堡礁的白化讓人心痛,看來地球暖化,氣候變遷,加上人為汙染,第六次大滅絕已非危言聳聽。





































#/media/Datei:Green_Island_1.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}