

- 文/邱名鍾│台大昆蟲系博士|嘉義大學生物資源學系研究助理
在自然界中,掠食者在奪取獵物的資源前必須徹底解除目標的武裝。一部分的生物採用了最直接的方法,成為捕食者將獵物殺死;但也有另一部分走向了完全不同的策略,牠們成為寄生蟲而依附在其他生物身上。在取食的過程中,獵物仍然活著。這個特性讓許多寄生蟲展出獨特的生活方式,並因此活躍在人類的想像之中。鐵線蟲便是其中典型的例子。
直到如今,鐵線蟲的研究在學術界都不算熱門,但一般民眾卻也還不算陌生,特別是 2012 年「鐵線蟲入侵」這部電影在韓國熱賣之後。影片的靈感取自於鐵線蟲造成寄主行為改變的現象,也就是大家熟知的昆蟲跳水自殺。電影對鐵線蟲生物特性的考據做的很到位,當然所謂的考據是對鐵線蟲特性的描述,不是讓人也一併跳水這方面。

寄主的跳水自殺現象是鐵線蟲研究最熱門的部分,這源自於寄生蟲的生存策略。鐵線蟲在陸生昆蟲體內發育,但卻必需進入水中繁殖。牠們本身缺乏在陸地上移動的能力,因此只能藉由操縱寄主的行為來回到水中。這個現象在 20 世紀初就被觀察到,並在近年內經由一系列的實驗而證實。相關的研究可以參考之前發表在網路上的文章《目前學術界對鐵線蟲引導寄主跳水的研究現況》。
然而在行為影響吸引走了大多數的目光之後,其他的影響往往被忽略,包括理論上最容易被觀察到的寄主外形變化。部分的螳螂可以藉由外形來判斷是否受到鐵線蟲的感染,也間接的為日後激烈的行為影響做準備。
最早被發現的鐵線蟲寄主形態變化
在 20 世紀中葉的 1940 年代,人類才剛開始接受鐵線蟲不是馬鬃毛泡到水而變成的生物。這時已經有科學家發現受感染的寄主外形變的「怪怪的」。這個怪怪的特徵來自一對雌螽斯的生殖器附肢,長到了雄蟲的身上。
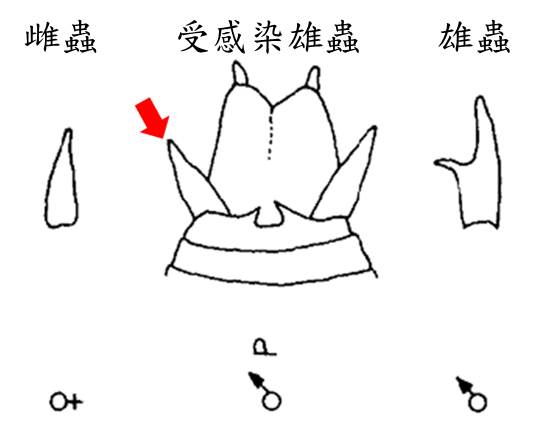
當時的科學家認為這是寄生蟲破壞性腺的結果,而這個推論來自於對哺乳類動物的了解。包括人類在內,許多哺乳類動物的性徵發育受到性腺分泌物 (如睪固酮)的調控。在部分的定義中,這些特徵被稱為第二性徵 (像人類的喉結或鬍鬚)。性腺被破壞後第二性徵的發育會一併受到干擾而出現「間性」的現象,使本來應該發育成雄性或雌性型態的構造產生畸型,或直接長成對方的樣子。
人類的疾病或內分泌失調中偶而會發現類似的現象,而這個印象便被投射到受感染的昆蟲身上。加上大多數寄生蟲造成的間性都伴隨著寄主性腺的破壞或萎縮,這個因果關係的推論看起來很有說服力。
然而隨著昆蟲賀爾蒙的研究,科學家發現之間的因果關係遠比想像中的複雜,甚至一度認為昆蟲可能根本不存在所謂的性荷爾蒙。但不論機制為何,寄主的性徵發育確實因感染而發生變化。並且隨著研究的累積,越來越多的寄生蟲被發現會對寄主造成類似的影響。
但為什麼寄生蟲要針對性徵?

在大多數的情況下,性徵代表著雙性 (雌雄)生物在繁殖策略上的分化。這源自於配子(精卵)生成時不平衡的能量投資。雄性生物產生精子所消耗的能量遠少於卵,因此有大量的資源能投資在求偶上,像雄獨角仙頭上那根醒目的犄角,讓他在領域競爭中佔盡優勢並增加跟雌蟲的交配機會。然而雌蟲需要耗費大量能量在卵的生成,因此在其他構造的發育上則能省則省。
換句話說,性徵的發育是為了繁殖而準備,但繁殖的投資對兩類生物而言是個負擔。
- 一是幼期生物,因為還沒輪到他們,所以雌雄間的差別在許多幼期生物身上並不明顯,他們偏向耗費更多能量來維持生存。
- 而另一個則是寄生蟲,寄主對繁殖的投資對他們而言是一種浪費。
這個浪費來自於寄生蟲對資源的競爭。
在個體的層級上,對寄生蟲最有利的情況是把寄主的資源盡可能全部轉移到自己身上,只要留下足夠寄主存活的能量即可,繁殖便盡可能的被排除。當然寄主族群消失對寄生蟲本身也是致命的傷害,因此這個現象有些前提:
要不是寄生蟲離開後寄主生殖能力會恢復 (像部分受感染的魚類或螺);就是像鐵線蟲一樣,不僅只是要掠奪生殖資源而已,從一開始就沒打算讓寄主活著。
殭屍寄生蟲,除了奪舍還做了什麼?
生物個體終將凋零,族群卻能因繁殖而延續。但這些生殖能力已經被破壞的寄主還能算是「活著」嗎?
曾經有科學家用奪舍 (body snatcher)來形容這些寄生蟲。這個意思接近另一個更常被使用的形容詞:殭屍 (zombie)。這兩個用詞雖然普遍但隱含很大成分的抽象描述,暗示寄生蟲控制了寄主的心智。這種比喻能讓人很快了解發生的大致情況,但在科學研究中,我們需要更具體的描述來了解:寄生蟲到底接管了寄主的哪些層面?
失去生殖能力的寄主被排除在族群延續的成員之外。牠們不再將基因向下一代傳遞,自身帶有的遺傳資訊在個體凋零之後便消失在世界上。在演化的時間尺度上,這不過是一瞬間的事。因此從演化的觀點出發,牠們已經從原本的族群死亡。
但牠們的生命跡象仍然持續在運作。

有另一組基因仍然依賴著這個生命現象的運作而傳遞。兩組本來互相獨立的遺傳訊息這時走向了一個共同的目標:
延續寄生蟲的族群。
從生殖能力遭到破壞,一直到生命跡象完全消失的這段時間,這些寄主身上出現的特徵已經不只來自本身基因的表現,還有很大成分從寄生蟲的基因延伸而來。這並非意味著寄生蟲改變了寄主的基因,更多的是寄生蟲透過化學訊號將自己的基因表現在其他的生物身上。所以很多寄主身上出現的異常行為或是發育,都能從寄生蟲的適應上找到答案。
這類寄生蟲通常會在感染的初期就開始嘗試接管寄主的剩餘人生。在鐵線蟲的例子中,幼蟲以包囊形式進入螳螂的一個月後,就已經有超過半數的寄主身上開始出現這些發育異常的特徵,而這時的鐵線蟲發育,也才經過不到一半的時間。也就是我們在野外看到的螳螂,很早就被抽離原本的族群,為了另一群生物的延續而繼續「活著」。

鐵線蟲與螳螂
螳螂身上出現變異的性徵,便是有鐵線蟲開始介入寄主發育與干擾生殖能力的證據。在 2003 年,科學家便從一隻受感染的螳螂身上發現性徵發育受到干擾的現象。不過大規模的樣本比較一直等到 2015 年才因台灣斧螳的高度感染與明顯的雌雄形態差異才得以問世。

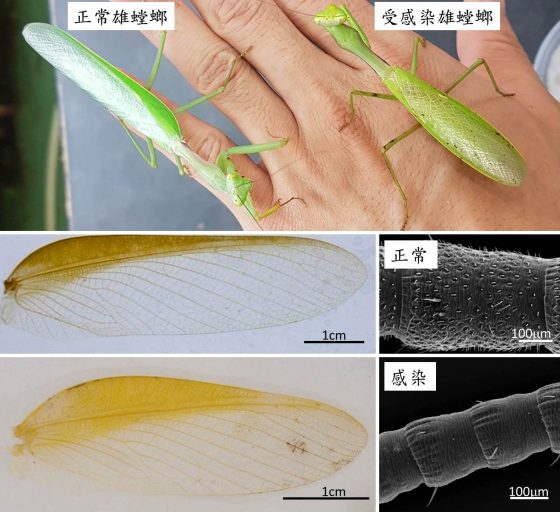
台灣斧螳 (Hierodula formosana)是台灣最普遍的螳螂之一,也是臺灣索鐵線蟲 (Chordodes formosanus)的主要寄主。每年的 6 月底到 8 月初是最容易在野外被觀察到的季節。台灣斧螳的雌雄成蟲有很明顯的差異:雄蟲細長,翅膀長且透明,飛行能力佳,觸角相對雌蟲長且粗,表面在顯微鏡下能看到密布的感覺器官;雌蟲體長與雄蟲相當但明顯較粗壯,翅膀厚實但幾乎沒有飛行能力,觸角相對較細且短。
這些特徵都是台灣斧螳的成蟲性徵,形態的差異主要源自於螳螂的交配行為。1970 年代末期科學家發現雌螳螂釋放費洛蒙吸引雄蟲交配的現象,而後來一系列的研究都指出許多種類的雄螳螂均具有較好的飛行能力(長而薄的翅膀)與嗅覺 (較長的觸角與觸角上高密度的感覺器官)。這些現象在台灣斧螳身上都能明顯發現,而也如同前文所說,這些性徵的發育將受到鐵線蟲感染而發生變化。

雄螳螂身上的雌性化特徵
螳螂的外形之所以能反映感染狀態,很大一部分便是來自這些產生變異的性徵。但有趣的是,這些變異大多只出現在雄蟲身上。在交配過程中,較長的翅膀與發達的嗅覺器官讓雄蟲能快速的偵測並移動到雌蟲附近,爭取交配機會。然而這些構造越發達,發育過程中所投資的能量就越多。
這些投資對鐵線蟲而言是多餘的,牠們只要寄主好好活著。
在比較上百個野外採集到的螳螂樣本之後,被感染的台灣斧螳的雄蟲被發現普遍出現翅膀變短與形狀改變的現象,這些變化使這些雄蟲的前翅形狀介於正常雌雄蟲之間。而觸角的變化更為明顯,除了長度變短之外,嗅覺器官的密度直接降低至與雌蟲相差無幾。而在解剖之後,進一步發現大部分個體的精巢 (睪丸) 不是不見就是萎縮到幾乎無法辨識的程度。
雄蟲的生殖構造差不多就是整組壞了,那雌蟲呢?
不論是前翅的形狀或是觸角上的感覺器官,雌螳螂都沒有因感染而受到明顯的影響。雖然大部分的雌蟲感染後體型變小,但外形基本上沒有太大的改變。解剖開後的卵巢形態雖然沒有明顯變化,然而只有極少數的個體能找到少量的成熟卵。相對於沒有受到感染的雌螳螂大多能在體內找到幾十到近百顆的卵,這些雌蟲雖然外形沒有受到影響,但繁殖能力可能也已經消失。
在形態變化之後
不論雌雄螳螂,牠們的繁殖能力都在鐵線蟲的感染後受到破壞。這些螳螂不再對自身的族群延續存在貢獻,在演化的過程中,便相對不容易發展出對抗寄生蟲的能力。這也可能讓鐵線蟲得以發展出少見的強烈影響:誘導寄主跳水,並回到水中繁殖。
這段寄生關係在鐵線蟲回到水中後結束。失去繁殖能力的螳螂,也大多在數小時到數天內死亡。然而目前的理解還不完全,對我們來說研究才剛開始。
鐵線蟲造成的性徵變化引發出一系列的問題,包含昆蟲性徵分化的機制。在螳螂的發育過程中,雌雄蟲的差異在剛孵化的若蟲身上並不明顯,不同的特徵要在特定的發育齡期才會開始分化。部分的性徵因感染而變化,但也有部分的特徵完全不受寄生蟲影響。有的部分一旦開始分化後便不再回頭,然而也有些是可逆的。這代表螳螂的性徵分化同時受到多個不同的生理機制調控,鐵線蟲只影響了其中一小部分。

然而,螳螂在受到感染之後真的只能任憑擺佈嗎?寄生蟲與寄主之間的軍備競賽從來沒有停止過。數學模型的預測一致的認為大多數種類的寄生蟲都應該發展出徹底「閹割」寄主的能力,然而實際案例卻大都辦不到。而在以往的認知中,螳螂釋放鐵線蟲後大多僅能存活數小時到 1 天,但越來越多的例子顯示部分的個體能存活更長的時間,甚至在蟋蟀的案例中出現產卵能力回復的現象。
還有很多現象等著進一步的研究。在這類研究中,我們往往不是在解決問題,而是在探討有多少問題還沒被發掘。近代對鐵線蟲了解的突破,大多都起始於對過往研究的質疑。目前有許多新發現的現象無法在現有理論下被完美的解釋,這些都是推動鐵線蟲下一步研究的契機。
延伸閱讀
- Chiu, M. C., C. G. Huang, W. J. Wu, and S. F. Shiao. 2015. Morphological allometry and intersexuality in horsehair-worm-infected mantids, Hierodula formosana (Mantodea: Mantidae). Parasitology 142: 1130-1142.
參考文獻
- Hurd, H. 2009. Evolutionary drivers of parasite-induced changes in insect life-history traits: From theory to underlying mechanisms. Advances in Parasitology 68: 85–110.
- Lafferty, K. D., and A. M. Kuris. 2009. Parasitic castration: the evolution and ecology of body snatchers. Trends in Parasitology 25: 564–572.
- Robinson, M. H., and B. Robinson. 1979. By dawn’s early light: Matutinal mating and sex attractants in a neotropical mantid. Science 205: 825–827.
- Roy, R. 2003. Répartition, biologie et variabilité de Tarachodella monticola Giglio-Tos, 1917 (Dictyoptera, Mantodea, Tarachodidae). Bulletin de la Societe Entomologique
- Wülker, W. 1964. Parasite-induced changes of internal and external sex characters in insects. Experimental Parasitology 15: 561–597.






































