Boto, E., N. Holmes, J. Leggett, G. Roberts, V. Shah, S. S. Meyer, L. D. Munoz, K. J. Mullinger, T. M. Tierney, S. Bestmann, G. R. Barnes, R. Bowtell and M. J. Brookes (2018). “Moving magnetoencephalography towards real-world applications with a wearable system.” Nature.
Cohen, D. (1968). “Magnetoencephalography: evidence of magnetic fields produced by alpha-rhythm currents.” Science161(3843): 784-786.
Cohen, D. (1972). “Magnetoencephalography: detection of the brain’s electrical activity with a superconducting magnetometer.” Science175(4022): 664-666.
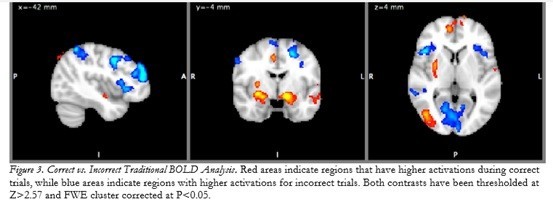
有趣的是,這份實驗也針對受試者判斷正確與判斷錯誤時的大腦變化進行比較,結果發現,當受試者判斷正確時,他們的大腦神經變化,會顯著地活化處理高階視覺反應的MT和側枕葉皮質(LOC) ,以及關於動作控制和處理的蒼白球(globus pallidus)和殼核(putamen) [7],還有牽涉到自我獎勵的額葉極區(frontal pole regions),且這些腦區的啟動,是不需要經過意識控制的!也就是說,他們的大腦會很自動地從辨認出球路切換到準備揮棒的模式裡,不需要經過大腦思考,這也是為什麼大聯盟傳奇名將Yogi Berra會說:「打者是不能一邊思考,一邊好好地打擊的(you can’t think and hit at the same time)。」。
Sherwin, J.Muraskin, & P.Sajda(2012)You can’t think and hit at the same time: neural correlates of baseball pitch classification. Front Neurosci, 6: p. 177.
Muraskin, J.Sherwin,& P.Sajda(2013)A System for Measuring the Neural Correlates of Baseball Pitch Recognition and Its Potential Use in Scouting and Player Development. Sports Analytics Conference.
Machielsen, W.C., et al., FMRI of visual encoding: reproducibility of activation. Hum Brain Mapp, 2000. 9(3): p. 156-64. 17.
Grill-Spector, K., et al., The lateral occipital complex and its role in object recognition. Vision Res, 2001. 41(10-11): p. 1409-22. 19. Hyvarinen, J., et al., Early visual deprivation alters modality of neuronal responses in area 19 of monkey cortex. Neurosci Lett, 1981. 26(3): p. 239-43
Kaas, J.H., Theories of visual cortex organization in primates: Areas of the third level. Extrageniculostriate Mechanisms Underlying VisuallyGuided Orientation Behavior, 1996. 112: p. 213-221. 18.
Tootell, R.B.H., et al., Functional-Analysis of Human Mt and Related Visual Cortical Areas Using Magnetic-Resonance-Imaging. Journal of Neuroscience, 1995. 15(4): p. 3215-3230. 21.
Marchand, W.R., et al., Putamen coactivation during motor task execution. Neuroreport, 2008. 19(9): p. 957-60.
Euston, D.R., et al., The role of medial prefrontal cortex in memory and decision making. Neuron, 2012. 76(6): p. 1057-70.
Goldberg, I.I., et al., When the brain loses its self: Prefrontal inactivation during sensorimotor processing. Neuron, 2006. 50(2): p. 329-339.
Philiastides, M.G., et al., EEG-informed fMRI reveals spatiotemporal characteristics of perceptual decision making. Journal of Neuroscience, 2007. 27(48): p. 13082-13091.
Ryu, J.Kim, A.Ali, W.Kim & S.J. Radlo (2015) Changes in baseball batters’ brain activity with increased pitch choice. Journal of Integrative Neuroscience, Vol. 14, No. 3369–381.