- 文/約翰.米勒 John H. Miller|卡內基梅隆大學(Carnegie Mellon University)社會和決策科學系的經濟學暨社會科學教授,也是聖塔菲研究所的外聘教師。現居美國賓州匹茲堡。
水和石頭在受到外力時選擇的路徑看起來很巧妙(雖然其實是受重力的影響)能向我們證明聰明並不受限於有智慧的生物。當我們回過頭來檢視生物時,這個主題還會變得更加有趣,因為生物做抉擇的競技場,和我們珍視的事物更是貼近。荷蘭科學家安東尼.范.雷文霍克(Antonie van Leeuwenhoek)把顯微鏡改良到能觀察單細胞生物之後,他注意到細胞以明顯有目標的方式移動。
往後數百年間,許多研究人員又更進一步發現某些類型的細胞和生物體會根據環境裡的化學信號引導自己的運動。
趨化性:生物隨化學信號改變運動方向
這種現象統稱為化學趨向性(chemotaxis),簡稱趨化性。我們可以從一種行動緩慢的細菌——大腸桿菌,來了解趨化性的運作方式。大腸桿菌的外表有好幾根半剛性的螺旋狀鞭毛。這些鞭毛各自附著於一個化學發動機,受其驅使並分別朝向正、反向轉動。當鞭毛以逆時鐘方向轉動,它們就全部排列整齊成如同一個軟木塞開瓶器的螺旋鑽,開始推動細菌,沿著筆直路徑前進。順時鐘方向轉動則會拆開螺旋,於是鞭毛就朝四面八方揮舞,而細菌開始任意翻滾。我們觀察到這種細菌的運動會在任意翻滾和筆直前行之間來回交替。
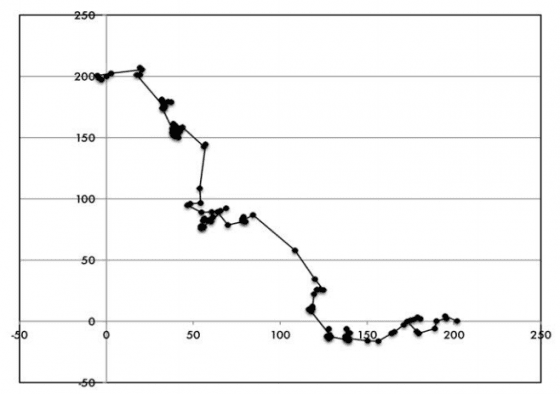
儘管這些行為看起來很有限,卻也足以讓細菌前往它需要去的地方。假定我們滴一滴化學物質到細菌的世界中。那滴物質會開始緩慢擴散,形成一種化學梯度,滴劑滴落的位置濃度很高,距離定點越遠,濃度也越低。假如我們滴進去的化學物質是糖之類的滋養糧食。一旦滴劑開始擴散,細菌會表現出一些有趣的行為。儘管細菌定然交替進行著筆直前進和翻滾的運動,但兩種動作花的時間已經開始依照所面朝的方向而改變。當細菌朝著滴劑移動,它往往花較長時間筆直前行,翻滾的時間則就較短。
當它的走向遠離滴劑,細菌則往往花較長時間翻滾。這類行為大體會讓細菌逐步攀登較高的化學梯度(圖 6.1)。相同道理,倘若化學物質是細菌想避開的東西,那麼當細菌的走向對準物質源頭,它就會傾向於較常翻滾;若走向遠離,它就較少翻滾,細菌也能因此脫離濃度較高的範圍。
我們知道,細菌完全沒有神經元或其他可以構成腦子的明顯部位,然而它卻有辦法朝向好東西靠近,並遠離壞東西。案例中,分子替代了腦子。儘管明確的分子機制和化學反應說起來有點複雜(起碼對經濟學家來講是複雜了點),細菌總歸就是由一套複雜系統掌控,而這套系統則是由依循化學規則彼此互動的分子所構成。
分子與受體結合,觸動陣級聯反應
當兩個分子相遇,當中的一個或兩個都可能出現變化。這種變化和分子實際外形息息相關,而且科學界投入很大部分的心力,專注探究蛋白質鏈如何摺出種種分子結構。偶爾會有某種分子和另一種分子「匹配」,使受體分子有所改變。有時分子則能相互增補或移除化學基,約略於開啟或關閉一種化學開關。一個分子持著這些核心能力,再攙入幾十億年份的演化作用,調節出種種不同回饋迴路和經歷一連串的蛻變歷程,最後才得以出現一些相當精妙的行為。
簡單的趨化性過程大致如下所述。當我們把一滴化學物質擺進環境,滴劑中數量龐大的分子(為了方便,就說有 6×1023 個左右)便立刻開始擴散。一段時間之後,這種擴散作用會形成一種梯度,當我們離開原來那滴物質愈遠,分子數量也愈少。
細菌在它的世界四處遊走,一邊也會遇見這些分子。細菌胞膜外側有受體,能輕易地與分子黏合——常見的比喻就是把分子想成鑰匙,能插進上頭有像是鎖的受體並把它開啟。當一個分子和受體結合,它就會觸動細胞內部,觸動陣陣級聯反應(cascade),並促使細菌表現出兩種重要的行為。
當細菌遇到一種它討厭的分子與受體結合時,受體因此觸發的第一種反應就是向細胞內部釋出一群新分子——為避免混淆,我們就稱後面這群新分子為信號。信號會促使細菌表現一陣陣級聯反應,把初始信號傳播出去,最後生成一種為時短暫的新信號,從而逆轉鞭毛發動機。
一般狀況中,這些發動機都朝逆時鐘方向轉動(約每分鐘六千轉),也就是筆直向前泳動,但當發動機逆轉時,鞭毛就全都斜向歪扭,而細菌開始翻滾。短效期信號引發翻滾後,發動機很快的便恢復常態旋轉,細菌也回歸正道。倘若和外表受體結合的並不是令它討厭的分子,而是種能誘引它的物質,那麼,這類逆轉信號就會比較少,細菌也就較少翻滾。所以,分子誘發的方式產生了一種適應力強的行為:當細菌身處它喜歡的物質附近,它往往就會筆直前行,一旦遇上了討厭物質,它就會改變方向。
第二種與外界分子結合的行為,牽涉外表受體本身的回饋迴路之一。外界分子與受體的結合和受體的敏感度息息相關,這也因此和某些內在變化的成因有緊密關聯。隨著結合現象愈頻繁,受體本身對外界分子的敏感度就會降低,其實也就是其適應了外界的平均化學物質含量——並延續一分鐘左右的短暫期間。當它喜愛的分子和受體結合,立刻(如前面討論)會減少翻滾次數,同時促使受體降低對喜愛物質的敏感程度,並延長時間。
倘若往後的幾分鐘內,細菌遇上了相仿濃度的誘引物質,它的翻滾頻率並不會縮減,而是依然保持常態。這種回饋機制讓細菌得以「記住」短暫過往。所以,倘若它發現自己身處片刻之前的相同處境,就會回復自己(偶爾翻滾)的常態行為,唯有當它感測出有別於短暫過往的改變,才會再度改變這個模式。一旦細菌發現自己身處與過往相似的情況時,這種記憶就會誘發細菌的探索行為。

前面我們專注探討細菌遇上喜愛或討厭物質時的狀況。另一方面,倘若它同時遇上了這兩種物質,又會發生什麼事?此時,細菌必須評估喜愛物質的潛在效益和討厭物質的潛在損失,並權衡取捨做出決定。這道問題最早在一八八八年由德國萊比錫大學的植物學教授威廉.普費弗(Wilhelm Pfeffer)率先探究,他發現,結果取決於兩種物質的相對濃度——倘若喜愛物質的濃度勝過討厭物質的濃度,則細菌就會向前行進,反之則遠離。
所以細菌尋覓好東西、避開壞東西的決策程序,也可以在兩種物質同時出現時做出取捨,細菌因此也擁有一套好惡偏私的標準。

本文摘自《直視全貌:穿越過度簡化的迷障,從複雜理論探索科學、商業與社會文化的新視角》,臉譜出版。