J編按:張鋒和其他兩位科學家伊曼紐.夏彭提耶(Emmanuelle Charpentier)、珍妮佛.道納(Jennifer A. Doudna),於 2016 年共同獲得第二屆唐獎的肯定。
文/國立臺灣大學醫學院生理所林世青專任研究助理
麻省理工學院的張鋒(Zhang Feng)教授憑藉其發展的 CRISPR(Clustered, Regularly Interspaced, Short Palindromic Repeats)/Cas9(CRISPR-Associated Protein 9)系統,年僅32歲即榮獲2013年《自然》雜誌評選之年度新聞人物首位,並獲得「DNA編輯大師」之稱號(“365 days: Nature’s 10,” 2013)。

究竟這令人折服的基因編輯技術的發展過程為何呢?西元 1987年時,科學家在細菌內發現一種特殊核酸內切酶,命名為 CRISPR/Cas9,其會辨認外來的 DNA並加以切割降解,被認為是細菌用以抵抗病毒感染的防禦機制(Ishino, Shinagawa, Makino, Amemura, & Nakata, 1987; Bhaya, Davison, & Barrangou, 2011),若能在病毒感染的第一時間內就將其 DNA降解,即可有效阻止病毒複製。
而到了2012年時,科學家解開了 Cas9對於目標 DNA的辨認機制,並成功藉由人工方式修改其專一性,讓整個技術有了飛躍性的突破。其發現 Cas9會與其導引 RNA(small-guiding RNA)結合後得到辨認目標序列的能力,從而結合並切割目標 DNA,科學家只需要修改導引 RNA的序列,即可改變 Cas9的專一性,命令其轉而裁切另一不同序列的 DNA(Jiang, Bikard, Cox, Zhang, & Marraffini, 2013; Jinek et al., 2012)。過去使用的基因剔除技術:鋅指核酸酶(zinc-finger nuclease, ZFN)或 TALEN標靶基因剔除工具等,都必須經由繁複多步驟的基因工程,拼裝出能夠辨認目標序列的蛋白區位,才得以用來辨認並切割特定序列(Gaj, Gersbach, & Barbas, 2013)。相較之下,因合成特定序列 DNA或 RNA的技術較拼裝重組蛋白更為成熟且有效率,僅需數日便可得到特定序列產物,令 CRISPR/Cas9的實行上較 ZFN或 TALEN更為方便快速。
張鋒教授的重大貢獻之處,即在於將本來不起眼的 CRISPR / Cas9的細菌免疫系統改造成為一套簡單廉價的基因改造工具(Hsu, Lander, & Zhang, 2014),並發現這套系統可以被用於原核或真核細胞的基因編輯,使得我們可以很簡單地自行在實驗室編輯各類常見模式生物,包括酵母菌(DiCarlo et al., 2013)、線蟲、果蠅(Kondo & Ueda, 2013)、阿拉伯芥、斑馬魚、小鼠、大鼠、乃至人類細胞的基因(Sakuma, Nishikawa, Kume, Chayama, & Yamamoto, 2014)。
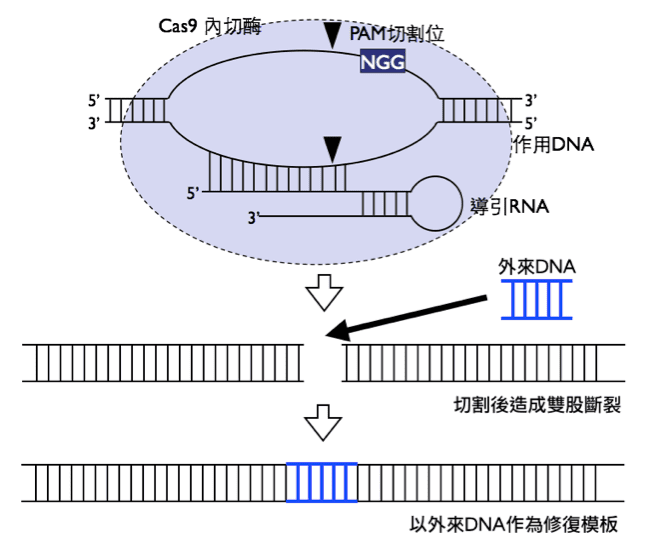
Cas9含有兩個內切酶作用位,當作用 DNA與特定序列之導引 RNA結合後,Cas9可同時切割該處雙股 DNA,造成 DNA斷裂並啟動細胞本身的修復機制,此時有一定機率會使用人工給予的外來模板修復該處,達到基因修飾目的,由此方法可建立基因修飾之各種模式生物,或往後應用於基因療法。
以遺傳學上常使用的模式生物:黑腹果蠅(Drosophila melanogaster)為例,根據全基因體定序分析結果,推算出果蠅約有超過15000個基因(Adams et al., 2000),過去的研究方法,必須使用例如EMS突變劑、跳躍子跳躍、費時的同源互換、繁複的 ZFN與 TALEN等技術,花費漫長時間才有可能得到目標基因的突變株果蠅,從而分析該基因的功能特性。

然而在發展Cas9技術後,只要得知目標基因序列,即可設計並合成出專門辨認該基因的導引 RNA表現載體,表現出導引 RNA後,Cas9即可快速專一地作用,直接剔除該基因,研究者僅花費很短時間與精力即可得到目標基因剔除的突變果蠅,效率為前人難以望其項背。
除了單純的基因剔除外,Cas9切割目標特定序列的特性還可以用於基因修飾,亦即先以 Cas9於目標基因處製造雙股斷裂,在有外來模板的情況下,細胞有一定機率使用外來模板修復斷裂處,從而將我們想置入的報導基因(Gratz et al., 2014),例如綠色螢光蛋白(green fluorescence protein, GFP)基因置入目標基因處,而得到帶有螢光蛋白篩選標記的突變果蠅。或是原先細胞基因即為帶有異常點突變的對偶基因,藉由外來模板給予正確序列的形式讓細胞得以修復該突變,令目標基因回復正常功能(Niu, Zhang, & Chen, 2014)。
由此可見,伴隨著不同的實驗設計,Cas9技術未來的發展無可限量。張鋒表示:「CRISPR / Cas9系統有助於幫助我們修正基因的微小突變。儘管只有少數人群攜帶這種致病突變,但這些突變對人類健康的影響則是災難性的(“365 days: Nature’s 10,” 2013)。」因為 CRISPR / Cas9在人類細胞基因剔除上已有顯著成果,故張鋒教授目前致力於利用該技術建立人類疾病相關基因之剔除資料庫,意味在理想狀態下,科學家可以根據該資料庫對人體任何器官中任何基因進行剔除或修正。而他自身最感興趣的部分即為利用該系統進行基因療法,治療如杭丁頓舞蹈症、囊狀纖維化或鐮刀性紅血球貧血症等因少數基因的微小突變造成之嚴重遺傳疾病,若能成功治癒,將是科學史以及醫療發展上的極大福音。未來的遺傳學與生物醫學研究,值得期待(Charpentier & Doudna, 2013)。
參考文獻:
- 365 days: Nature’s 10. (2013). Nature, 504(7480), 357-365. doi: 10.1038/504357a
- Adams, M. D., Celniker, S. E., Holt, R. A., Evans, C. A., Gocayne, J. D., Amanatides, P. G., … Venter, J. C. (2000). The genome sequence of Drosophila melanogaster. Science, 287(5461), 2185-2195.
- Bhaya, D., Davison, M., & Barrangou, R. (2011). CRISPR-Cas systems in bacteria and archaea: versatile small RNAs for adaptive defense and regulation. Annu Rev Genet, 45, 273-297. doi: 10.1146/annurev-genet-110410-132430
- Charpentier, E., & Doudna, J. A. (2013). Biotechnology: Rewriting a genome. Nature, 495(7439), 50-51.
- DiCarlo, J. E., Norville, J. E., Mali, P., Rios, X., Aach, J., & Church, G. M. (2013). Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res, 41(7), 4336-4343. doi: 10.1093/nar/gkt135
- Gaj, T., Gersbach, C. A., & Barbas, C. F., 3rd. (2013). ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol, 31(7), 397-405. doi: 10.1016/j.tibtech.2013.04.004
- Gratz, S. J., Ukken, F. P., Rubinstein, C. D., Thiede, G., Donohue, L. K., Cummings, A. M., & O’Connor-Giles, K. M. (2014). Highly specific and efficient CRISPR/Cas9-catalyzed homology-directed repair in Drosophila. Genetics, 196(4), 961-971. doi: 10.1534/genetics.113.160713
- Hsu, P. D., Lander, E. S., & Zhang, F. (2014). Development and applications of CRISPR-Cas9 for genome engineering. Cell, 157(6), 1262-1278. doi: 10.1016/j.cell.2014.05.010
- Ishino, Y., Shinagawa, H., Makino, K., Amemura, M., & Nakata, A. (1987). Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J Bacteriol, 169(12), 5429-5433.
- Jiang, W., Bikard, D., Cox, D., Zhang, F., & Marraffini, L. A. (2013). RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat Biotechnol, 31(3), 233-239. doi: 10.1038/nbt.2508
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., & Charpentier, E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.Science, 337(6096), 816-821. doi: 10.1126/science.1225829
- Kondo, S., & Ueda, R. (2013). Highly improved gene targeting by germline-specific Cas9 expression in Drosophila. Genetics, 195(3), 715-721. doi: 10.1534/genetics.113.156737
- Niu, J., Zhang, B., & Chen, H. (2014). Applications of TALENs and CRISPR/Cas9 in Human Cells and Their Potentials for Gene Therapy. Mol Biotechnol, 56(8), 681-688. doi: 10.1007/s12033-014-9771-z
- Sakuma, T., Nishikawa, A., Kume, S., Chayama, K., & Yamamoto, T. (2014). Multiplex genome engineering in human cells using all-in-one CRISPR/Cas9 vector system. Sci Rep, 4, 5400. doi: 10.1038/srep05400
本文原發表於科學Online-科技部高瞻自然科學教學資源平台,經授權轉載。
「科學Online」在科技部高瞻計畫指導下執行,為一具有指標性與權威性的科教網站,擁有以大學教授為主的專業責任編輯群,強調科學知識推廣的正確性,涵括豐富的自然科學實驗與講座影音、教學與學習資源、科技報導、科學圖鑑等,並結合App、Podcast與社群網站等媒體,提供科學教育多元且無地域界限的自主開放學習園地。




































《運動基因》立體封面72dpi.jpg)