尋找葉綠體內存在億萬年的蛋白質橋樑,解開光合作用之謎
本文轉載自中央研究院研之有物 ,泛科學為宣傳推廣執行單位
葉綠體的蛋白質橋樑
大家都知道光合作用是在葉綠體進行,但葉綠體有賴細胞輸入「蛋白質工人」才能正常運作。中研院分子生物研究所特聘研究員李秀敏與其團隊,花了七年的時間,找到能讓蛋白質穿越葉綠體外圍雙層膜的橋樑 TIC236,解開葉綠體運作的大謎團,更發現這套運輸系統從遠古細菌一直沿用到高等植物,是植物演化學的重大突破,論文於 2018 年 12 月登上《自然》 (Nature),並獲專文推薦。
踏入李秀敏 的實驗室,窗邊桌面是綠意盎然的植物,牆上貼著玉米品種演化和花草彩繪海報,拍照時,研究人員搬來幾株植物一同入鏡,這間研究室與植物很親密!問起為何投入葉綠體的研究?李秀敏想了想:「嗯,還真有點故事呢!」
李秀敏與她的研究團隊,由左到右為朱瓊枝、李秀敏、陳麗貞、陳奕霖。他們手上拿的就是本次研究的主角:阿拉伯芥和豌豆。 從小會幫植物取名字的小孩……長大唸了動物系? 她從小住在臺中眷村,孰悉巷弄裡的每一株植物,還會幫它們取名字。高中時成績很好,卻不想唸醫科,填志願時按照分數將臺大動物系填前面、植物系放後面。她笑著說:「小時候不知天高地厚,認為植物可以自己唸,動物比較不喜歡,請老師教,這樣就兩門都會。」
結果,唸動物系第一年,她就發現不對勁!解剖青蛙時,她覺得有些蛙骨沒用就扔了,沒想到每個骨頭都有名字;其他對動物有興趣的同學,對蛙骨卻如數家珍。雖然成績是班上第一,但她唸得很痛苦。
直到大二開始上普通植物學這門課,當課本發下……
我就有回到家的感覺,我有興趣的東西都在裡面!
李秀敏決定轉系,將動物系的課全退掉,選修植物系的課。她跑去找動物系主任簽名,主任皺著眉頭說:「你要簽切結書,如果沒法順利畢業,要自己負責!」
她又跑去找植物系的教授加選課程,教授也十分驚訝:「從來沒人從動物系轉到植物系,你確定嗎?」期末成績出爐,她得到最高分,終於在大三時,如願轉入植物系。
後來赴美留學,一開始她也不是研究植物,而是挑了酵母菌、藍綠菌和動物細胞的三個實驗室輪流實習,但內心始終無法滿足。
她回憶:「某天我在想,如果要花生命中最精華的四、五年寫一本博士論文,我會做酵母菌或動物嗎?不可能!一定是做植物。」後來,研究藍綠菌的老師建議她去一間研究葉綠體的實驗室,她的博士論文就在那間實驗室完成。
從此,她的人生就跟「葉綠體蛋白質運輸」這個題目,結下不解之緣。
葉綠體除了行光合作用,還有其他工作 說到葉綠體 ,李秀敏就像聊起老朋友般熱絡:「一般人對葉綠體的印象只是行光合作用,其實它還有許多功能,身世也很有趣。」
遠古時期,有顆單細胞動物吞下了藍綠菌 ,藍綠菌變成細胞內的葉綠體,從此演化出植物這個大家族。植物細胞更將細胞質的許多功能轉交給葉綠體執行。
李秀敏說,如果把植物細胞看成一座城市,細胞內的葉綠體就像是一間間「城市農園」,除了行光合作用製造養分,還要負責製造必需胺基酸、脂肪酸和荷爾蒙等物質。
農園,當然需要工人囉!「葉綠體農園」在運作時,需要許多具有特殊功能的蛋白質,但這些「蛋白質工人」,大部分都是由植物細胞的細胞核下令、在細胞質製造完成後,才送入葉綠體工作。
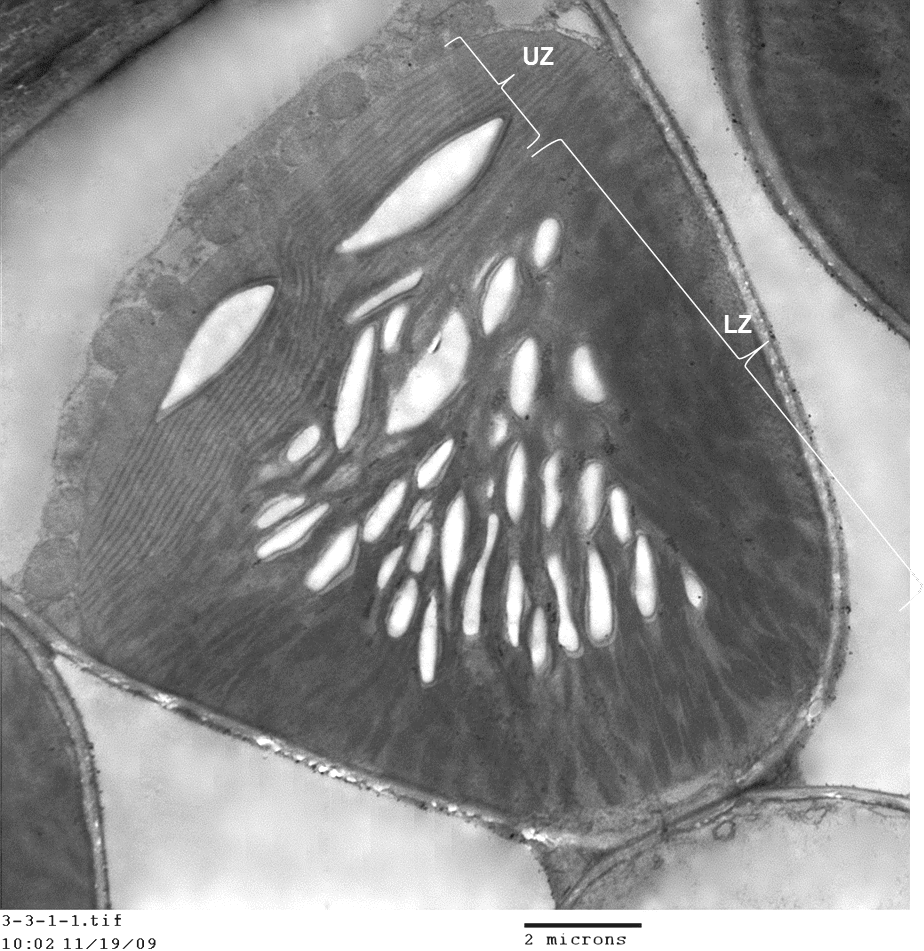
問題來了!葉綠體外表有外膜和內膜,就像兩道城牆,中間還隔著一道膜間隙,就像護城河。那麼,蛋白質工人到底是如何順利的進入葉綠體的呢?這就是「葉綠體蛋白質運輸」的謎題,困擾了科學家數十年。
葉綠體有外膜和內膜,就像內外兩道城牆,中間隔了膜間隙,就像城牆之間的護城河。過去科學家已經找到蛋白質進入外膜與內膜的轉運蛋白 (內外城門),並推測兩道城門應該有一座橋連接,讓蛋白質能順利跨越膜間隙 (護城河)。李秀敏團隊的貢獻就是找到這座橋:TIC236! 繞了一圈,回到最愛! 雖然她覺得葉綠體研究很有趣,但博士後為了拓展視野,她又拐了一個大彎,找了一個當紅的、研究植物光反應的實驗室。後來,她因緣際會飛去德國一個實驗室做研究,不料剛下飛機的第二天,騎腳踏車時摔斷了手!
她在醫院收到美國研究室傳來的植物光反應的實驗結果,心情很沮喪、根本不想看,就將紙翻面,沙沙沙寫下博士班沒完成的葉綠體題目……。
「又來了!你又忘記該追求自己的興趣。」她立刻寫信給博士後的老闆,放棄那個當紅的題目,回去研究最愛的葉綠體,從此再也不動搖。
「我現在常跟學生講,一定要追求自己的興趣,這樣才不會覺得累,可以一直做、一直做。」李秀敏微笑總結。
藏在葉綠體內外膜的謎團 回到這次的研究,李秀敏發現的葉綠體蛋白質橋樑,為什麼引起學界震撼?
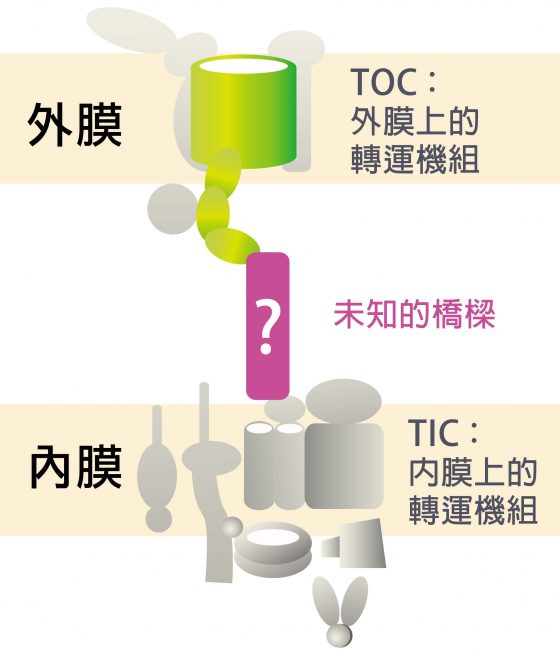
早期研究已經知道,要進入葉綠體的蛋白質會攜帶一段特殊信號,就像帶了「識別證」,內外膜上有轉運機組,就像膜上的城門,能夠辨識信號並讓蛋白質通過。歷經二十多年研究,科學家已陸續發現轉運機組的成員,並將外膜的轉運機組稱為 TOC ,內膜的轉運機組稱為 TIC。
但近年來,科學家陷入瓶頸:他們發現蛋白質會同時穿越葉綠體的內外膜,可是中間隔著寬闊的膜間隙 (護城河),蛋白質是怎麼穿越的?所以 TOC 和 TIC 這兩道「城門」之間,應該有一座「橋」相連,實驗上也支持這個看法。
怎麼證實的?科學家從豌豆苗分離出葉綠體,利用和膜上機組成員對應的抗體,將膜上的機組成員拉下來,看看能不能把內外膜機組成員一起拉下來。李秀敏解釋:「用抗體從外膜拉,TOC 和 TIC 會一起被拉下來,用抗體從內膜拉,兩者也一起被拉下來,所以知道彼此間一定有一座橋相連。」然而科學家始終找不到那座「橋」。
於是全世界有十幾個實驗室都嘗試想找到這座橋,李秀敏的實驗室則在 2018 年宣告破解這座神祕的橋樑:TIC236。
過去科學家已經找到內、外膜上的轉運機組的許多成員,實驗上也證實兩機組應該是相連,然而科學家始終找不到那座「橋」。 好運,是留給準備好的人 為什麼大家一直找不到,李秀敏的團隊卻能?因為他們突破了過去實驗的思考框架:不從葉綠體下手,而是研究白色體 !
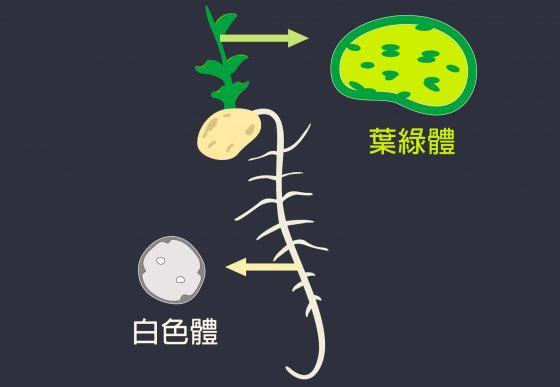
白色體和葉綠體,都是由植物細胞的色質體分化而來,所以結構很相似,只是在葉子為了行光合作用,變成含有葉綠素的葉綠體,在根部不行光合作用,變成不含葉綠素、無色的白色體,用來儲存養分。
豌豆葉片上的葉綠體與根部的白色體,兩者都是植物的色質體分化而來,所以結構很相似,一樣都具有內外膜的橋樑:TIC236。 過去科學家用葉綠體做實驗,分析葉綠體內成分。但葉綠體有許多跟光合作用有關的蛋白質,用質譜儀分析時,它們的量很大、訊號強,TIC236 因為很容易被水解、訊號弱,會被掩蓋。但李秀敏用白色體做實驗,沒有上面說的大量葉綠體的蛋白質干擾,TIC236 才有機會現身。
妙的是,當初李秀敏和團隊一開始研究白色體,也不是為了尋找內外膜之間的這座橋,而是想要知道白色體內外膜的轉運機組是否跟葉綠體一樣,想用在作物改良上。
她請團隊成員朱瓊枝取下豌豆根、分離出白色體,分析膜上的轉運機組成員,結果意外看見一個新面孔:TIC236!
李秀敏立刻上網搜尋,找到一篇論文提到大腸桿菌的雙層膜之間也有個作為橋樑的 TamB 蛋白,居然跟 TIC236 的序列相似,換句話說,TIC236 可能與 TamB 一樣具有「橋樑」的功能。「莫非 TIC236 就是葉綠體外、內膜間的橋樑?」李秀敏敏感的聯想。
她說:「好運是留給準備好的人。那天搜尋到它跟大腸桿菌的 TamB 橋樑蛋白很像時,我已經知道大概是什麼樣的故事,但要有本事去證明。 」
找到 TIC236 的闖關之路,比想像更艱辛 她先請團隊成員陳奕霖去研究:TIC236 基因是否在葉綠體裡也很重要?查資料庫後發現,TIC236 基因在阿拉伯芥 全株都有表現,一旦基因被破壞,植株就會死亡,但過去科學家並不清楚這個基因的作用。
阿拉伯芥的野生株 (左) 以及 TIC236 基因表現量減少的突變株 (右),突變株有發育不良或葉子缺刻等狀況,表示若缺少 TIC236 這座橋,葉綠體無法正常運作。 接下來,如果能證明 TIC236 和外膜通道蛋白 TOC75 真的相連,就能證明 TIC236 是連接內外膜轉運機組的橋樑了!陳奕霖用了一個很短的化學交聯劑 做實驗,它的兩端可以分別和不同的氨基酸相黏。如果 TIC236 和 TOC75 都被它黏住,表示兩者距離非常非常接近,應該是相連的。結果,實驗成功了!
但好事多磨!當時有個德國研究團隊已經發表論文,認為葉綠體外膜通道 TOC75 在演化過程中已翻轉了 180 度。李秀敏很詫異:「如果 TOC75 真的已翻轉,TIC236 就不可能和 TOC75 相連!」因此他們又繞了一大圈,花上兩年重複德國的實驗,終於證實他們是錯的。
蛋白質啊!你可不可以跑快一點 最後,怎麼讓大家「眼見為憑」:在 TIC236 基因表現量減少的突變株中,TOC 和TIC 轉運機組的複合體含量真的會減少?這要透過凝膠電泳並拍照。
方法是:通電後,各個蛋白質會按分子大小和性質等,分別在凝膠上移動,就像賽跑般有快 、有慢,藉此可將它們分開,一個個在凝膠上現形,就能一眼看出它們的多寡,又稱為跑膠。
一般蛋白質的分子量小,容易跑膠,但 TOC 和 TIC 轉運機組的複合體很大,在膠上跑不動。她說:「那時做這個真的快抓狂!那不是一般的膠,超難跑的!每個周末我都坐在研究室思考怎麼改進。」她和陳麗貞花了近兩年,終於完成這項艱鉅任務。
歷經七年的實驗抗戰,總算蒐集到充分證據,他們終於證明 TIC236 的確是葉綠體內、外膜之間的橋樑通道,並證實了這套運輸系統從最早的細菌一直保留至今,是植物演化學上的一大突破!論文獲登國際知名期刊《自然》(Nature),還得到編輯專文推薦。
李秀敏笑著解釋:「這個蛋白質很難做,我們有辦法找到,並證明它如同我們想像的,是從低等細菌一直保留到高等植物,是我們厲害的地方!」問到為什麼這次研究如此有創意,她頓了頓,認真的說:
這次的突破並非創新或天才想法,而是一步步、按部就班,並加上團隊的通力合作才能完成。
基於天生對植物的熱愛與長年的紮實研究,終於讓李秀敏發現這座葉綠體不可或缺的生命之橋。一直在這個問題上努力不懈的她,也宛如一座橋,引領我們探索植物生命的奧妙。
延伸閱讀
___________科學生 跟著課程進度每週更新科學文章並搭配測驗。來科學生 陪你一起唸科學!