

- 作者 / 蔡政修|台灣大學生命科學系、生態學與演化生物學研究所、動物博物館
相信生活在台灣的大多數人都會希望能有著花不完的千元大鈔可以自由的使用。身為古生物學家,除了一方面幻想著能有數不盡的千元大鈔可以讓我花費在野外挖掘、探索遠古生物的滅絕與生存奧秘之外,我也常常盯著千元大鈔上的帝雉(學名為:Syrmaticus mikado,俗名又常被稱為「黑長尾雉」),思考著帝雉在台灣的演化歷程到底是怎麼一回事,像是什麼時候來到台灣、又是如何演化成只有在台灣才能親眼觀察到野外族群的特有種呢?

餐桌上的烤雞、公園裡的麻雀,都是貨真價實的恐龍!
思索著這樣的古生物問題,或許和大多數人對於古生物研究的想像有點出入——在「侏羅紀公園」深遠的影響下,似乎只有研究暴龍、三角龍等主要出沒在中生代的恐龍們才能算是真正的古生物學研究一樣。但這也就是古生物、或其他基礎研究發現新知的樂趣一樣,藉由 1993 年幾乎風靡了全世界的「侏羅紀公園」電影,讓更多人、經費投入到古生物學的研究工作,到了現在,我在上課時所使用的恐龍教科書(由 S. Brusatte 所撰寫的《Dinosaur Paleobiology》)裡面清楚的寫著恐龍的定義為:
滅絕的三角龍和現生的麻雀最近的共同祖先、以及這一個共同祖先所有的後代都是恐龍。
換句話說,我們日常生活中會見到的麻雀、餐桌上美味的雞、鴨料理等,都是貨真價實的飛天恐龍和恐龍美食。也就是說,當我們翻開台灣生物多樣性目前的記載,台灣現今有超過 600 種以上的鳥類,也就可以說是台灣有 600 多種的恐龍——但更令人興奮的,在如此高度的恐龍多樣性裡,台灣有著 30 種左右的特有種恐龍!登上台灣千元大鈔的帝雉基本上就是台灣恐龍的代表物種之一。

具備了這樣的認知後,台灣目前的古生物研究裡即使基本上沒有暴露在地表面的中生代沉積地層能讓我們尋找、挖掘幾乎每個人都能琅琅上口的暴龍、迅猛龍、三角龍等在中生代、新生代交界的 6 千 6 百萬年前就完全滅絕的「非鳥類恐龍(non-avian dinosaurs)」,但進入新生代後、恐龍的演化仍是相當蓬勃,在這一個當下,我們稱為「鳥類」的恐龍們,在基礎的生物多樣性分類中,物種數也有 1 萬種左右、這數字大約是現生哺乳動物的兩倍!
台灣也有恐龍化石,只是沒有人去尋找它們
在這樣的知識架構下,台灣也當然是一定要有、會有新生代的恐龍化石,畢竟當下我們眼前超過 600 種以上的恐龍、也有 30 種的特有種恐龍都不會是突然間蹦出來、而是應該有久遠的起源、演化歷程。就好像是一開始所提到的、在台灣的大家所熟知的特有種帝雉,不會是一開始來到台灣就是特有種,而一定是由目前仍未知的「祖先」來到台灣後,經歷了獨特的生存歷程、一代又一代的演變,搖身一變成為了在全世界其他地區都看不到、只有在台灣才能一睹風采的台灣特有種。
出人意料的是,翻開台灣從日治時期由早期日本籍的古生物學家們、像是著名的早坂一郎開始在台灣有系統的探索、研究化石到現在,卻是連一件正式的恐龍化石紀錄也沒有、就是高掛著一顆大鴨蛋。但這也就是科學研究、思維的重要性,即使長期以來在台灣都沒有正式的恐龍化石紀錄,但如文章前面所陳述的,台灣目前有這麼多恐龍、尤其是有 30 種左右的特有種恐龍,我們的腳底下一定會有恐龍化石。當然不是很容易尋找,但他們就是仍埋藏、沈睡在某處的沈積岩裡,等著我們願意有系統的投入相對應的資源、心力去探索——畢竟化石不會自己跑來我們面前說著:請研究我們吧。

我 2018 年要從日本搬回台灣、在台灣大學生命科學系建置起古脊椎動物研究的實驗室前,我的研究方向其中之一就包含了要找尋、研究台灣至今仍沒有被知道、重視過的恐龍化石。也不意外的,當我真的回來後,幾乎每一個人都會問我說:「台灣真的有恐龍化石可以研究嗎?」之類的。沒有確切的證據之前,解釋著從古生物學的推論來判斷一定會有恐龍化石的相關說明後,也沒有太多的用處,畢竟大多數人就是想要看到真的實際有從台灣發現的恐龍化石。
默默耕耘的台灣化石獵人們
台灣看似不大、但如果真的到了野外,會感受到自己一個人能走到的野外地點、尋找到的化石標本極度有限。不過,台灣的古生物學其實從日治時期開始除了有日本籍如早坂一郎等古生物學家在撰寫相關的研究文章之外,其實早坂一郎也培養出了台灣當地的「化石獵人」,像是被稱為化石爺爺的陳春木。二次大戰結束後,早坂一郎和其他的研究人員陸陸續續回日本後,陳春木仍是默默的在台南的野外地區收集相關的化石標本,也因為這樣影響了一些當地民眾,帶起了更多人一起成為化石獵人。
陳春木在我還沒有機會遇到之前的 2002 年過世。但我到台南一邊在野外找化石之餘,也一邊和後繼的化石獵人聯絡、討論台灣古生物學研究的狀況和可能性。這段期間我也連絡上侯立仁——一名在台南相當著名、但低調的化石收藏家。我不時的表達出我正在尋找台灣的恐龍化石(也就是「鳥類」化石),在多次的拜訪、討論後,侯立仁拿出了一件他收藏了超過 20 年的化石標本——很清楚的就是恐龍腳部跗跖骨的一小段。
在我超級興奮之際,來來回回的討論過後,侯立仁也願意將這一件得來不易、在台灣的古生物研究領域中是極度稀少的化石標本捐贈給我,讓此化石標本進到我所任職的台灣大學生命科學系的古脊椎動物演化及多樣性實驗室裡的典藏系統永久保存和後續的研究!
有了侯立仁所捐贈的化石標本,現在,台灣的恐龍化石紀錄不再是一片空白!此發現也已經正式的刊登在國際間鳥類學的研究期刊中(Journal of Ornithology)。
台灣第一件恐龍化石,與帝雉、藍腹鷴同為雉科成員
這一件新生代的鳥類恐龍約略生存在 40 萬到 80 萬年前、出現的地點為目前的台南。化石標本即使沒有很完整,只有保存了左腳跗跖骨(確切的骨骼結構名稱為:tarsometatarsus)的遠端一小段,但主要的關節面和形態都保存的相當完整,足夠讓我們能從事後續的形態分析和研究。
跗跖骨的第三滑車的上部接合面是不對稱的形態,是可以辨識為雞形目成員的關鍵特徵。再更近一步的觀察手上所握有的這一個化石標本,跗跖骨的第二和第三,或是第三和第四滑車結構的間距都不大,加上第二滑車的構造有向足底面彎的形態,清楚的指出這一件只有保存了跗跖骨遠端約 2 公分長度的化石標本隸屬於雉科的物種。
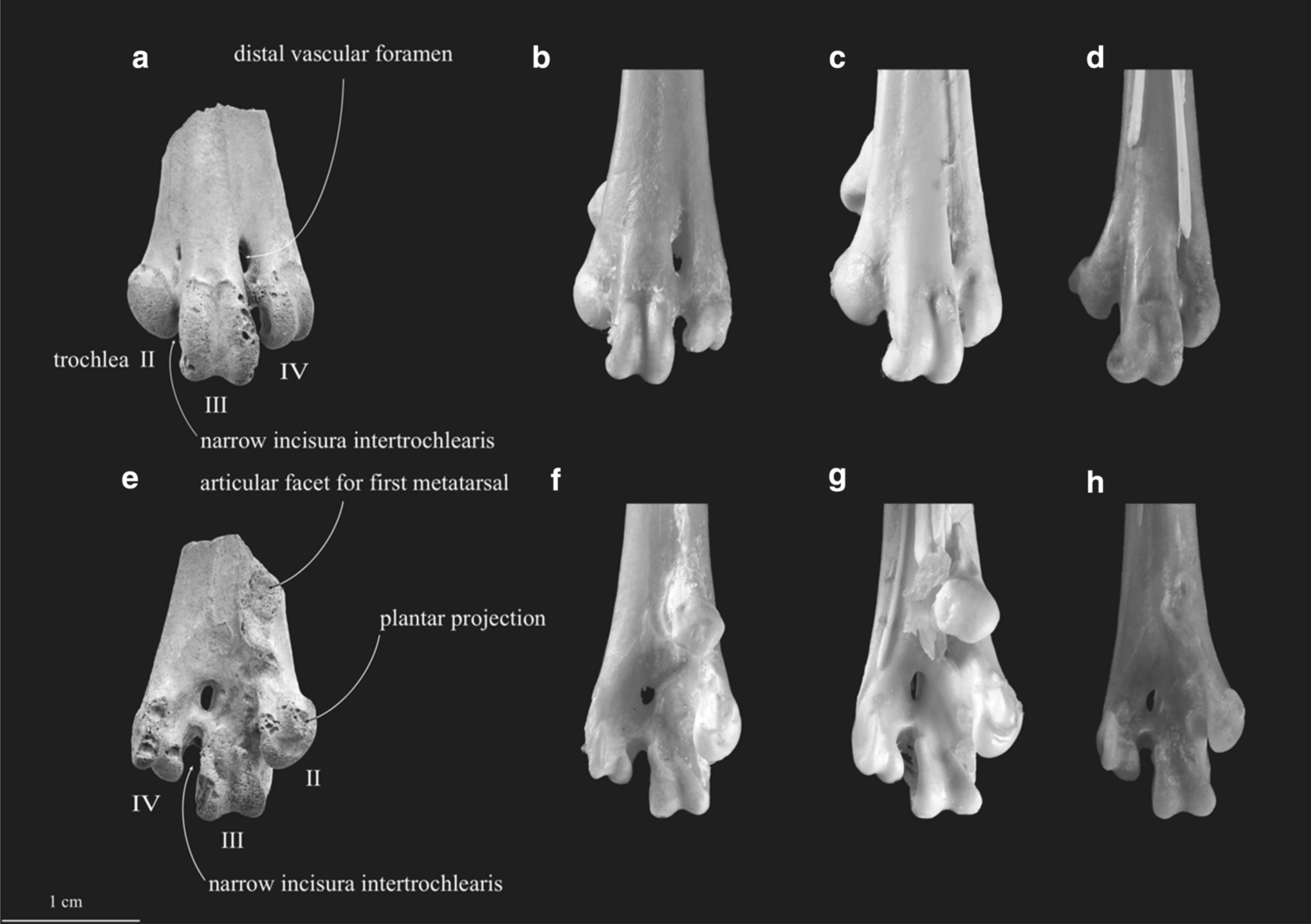
提到「雉」這一個字,或許有人已經聯想到了我們一開始所提到的「帝雉」,沒錯,帝雉就是台灣雉科成員的代表性物種之一。更令人著迷的是,台灣的雉科成員目前有 7 種原生種,但有超過一半的物種(4 種)是特有種!換句話說,台灣雉科的鳥類恐龍類群中,形成特有種的比例相當高,有:帝雉、藍腹鷴、台灣竹雞、台灣山鷓鴣(環頸雉被認定為是台灣的特有亞種,還沒有達到「特有種」的地位)。

看到確切的化石標本時,大多數的人或許都需要睜大眼睛來看,因為保存的長度是 2 公分多、寬度也只有 1 公分多一點。但這第二、第三、第四滑車集結所形成的跗跖骨寬度能有一公分多,其實在鳥類恐龍裡是很大的跗跖骨,在台灣的雉科物種裡能達到這樣大小的也只有:帝雉、藍腹鷴、和環頸雉。換句話說,台灣第一件的鳥類恐龍化石即使沒有很完整,但也能近乎奇蹟似的鎖定到帝雉、藍腹鷴、或環頸雉等大型的雉科成員。
跗跖骨化石是個起點,未來還有豐富的證據等待被挖掘
化石標本看起來不完整、形態鑑定上似乎也很有限,或許會讓很多人對於台灣第一件恐龍化石感到失望。不過,換個角度想,這也就是從事研究、探索未知的樂趣,一開始的發現或許看似不起眼,但那從零到一的突破,其實是有機會能引起無限的可能性、和後續更大的研究力道。就好像帝雉在 1906 年被發現、命名時也只有兩根尾羽、始祖鳥在 1861 年第一次公開發表時也只有一根羽毛。又或是歷史上第一件有正式學名的中生代非鳥類恐龍的命名可以回到 1763 年、也只有腳部股骨遠端的一小段,而且被取名為或許有點不雅、但令人難忘的:人陰囊(學名:Scrotum humanum)!目前這一個學名已經被Megalosaurus bucklandii(可翻成:巴克蘭巨龍)所取代。

這樣的起點不僅沒有減損其價制,反而造就了更迷人的故事、引發了更多人的幻想,從而有了更多的研究成果和大家的重視。或許還是有人會覺得這一個發現不是中生代的非鳥類恐龍,不夠酷炫之類的想法——也就更因為是古生物學家,我也才更加寄望能藉由提供正確古生物學知識。如文章一開頭所提到的:鳥類就是貨真價實的恐龍,來顛覆台灣長期以來幾乎沒有古生物學家能提供這樣顛覆世界觀的思維給大家來重新看待我們眼前的世界——台灣就是有恐龍化石可以挖掘、研究,也需要我們投入更多的心思、資源,才能利用台灣的恐龍化石研究跟世界接軌。
參考資料
- Tsai CH and Mayr G. 2021. A phasianid bird from the Pleistocene of Tainan: the very first avian fossil from Taiwan. Journal of Ornithology doi:10.1007/s10336-021-01886-w








































{kind=link}