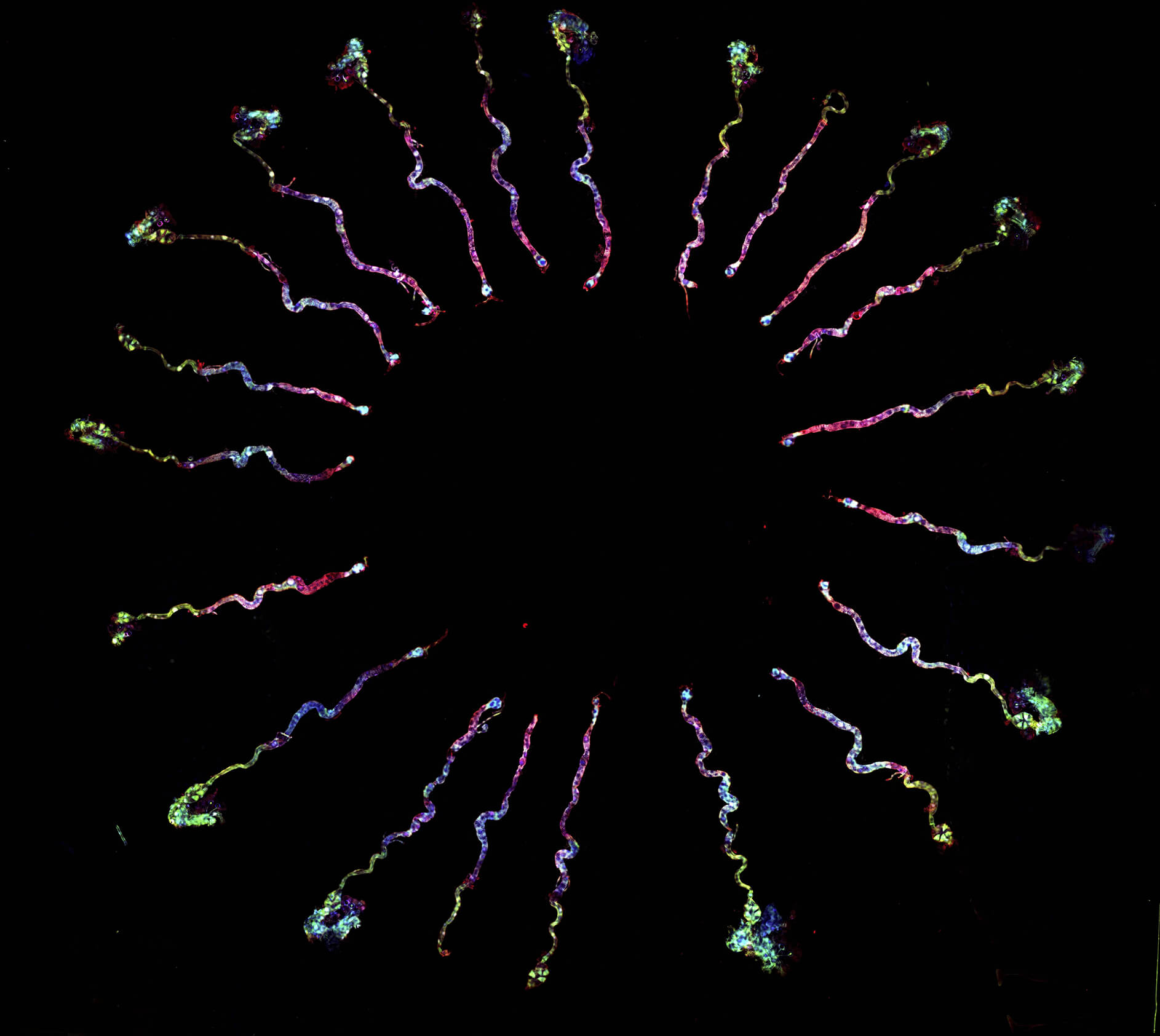
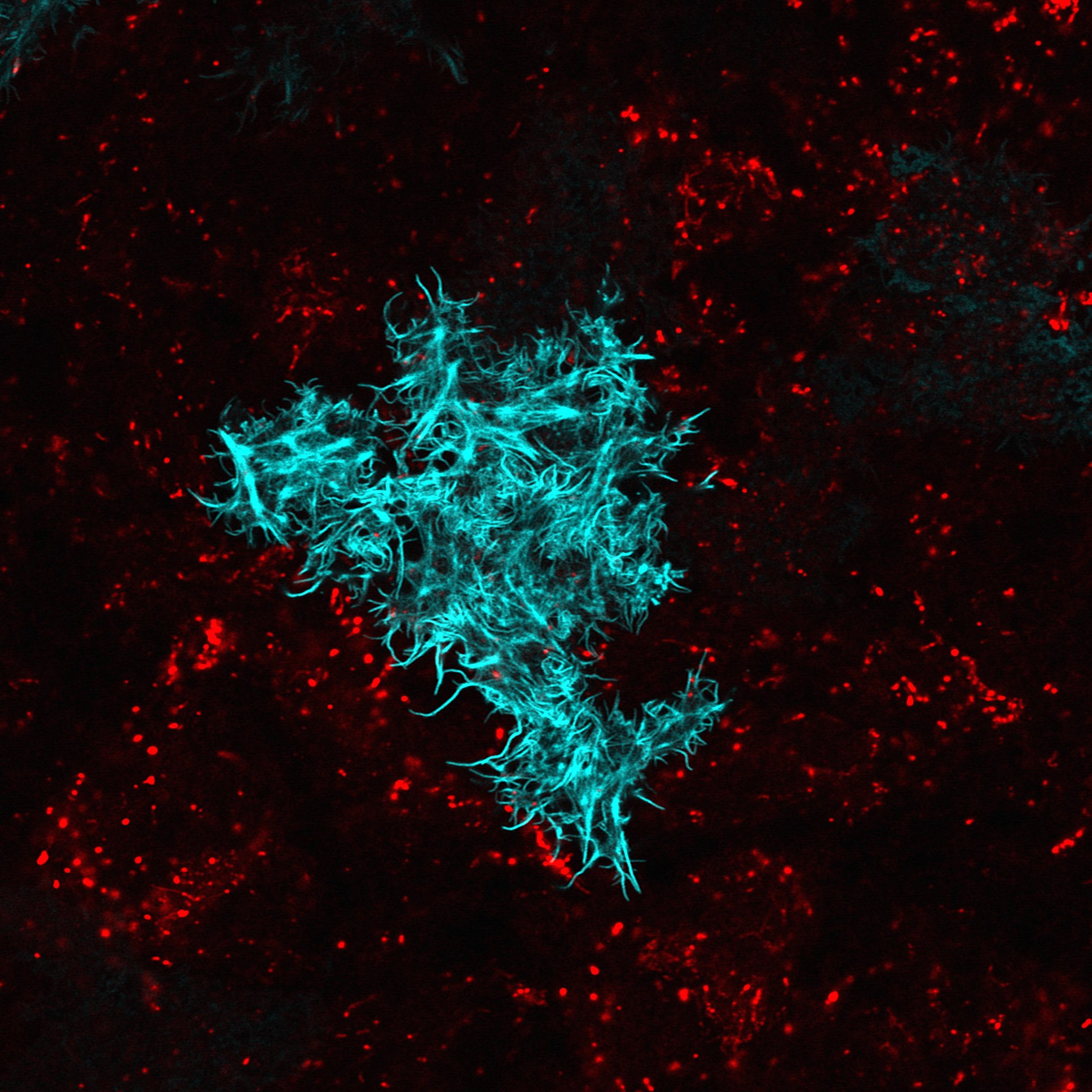
有頭有尾巴,長得像蝌蚪,大概是大家對精子的印象,不過某些精子的形象或許有點驚人。比方說有些果蠅(Drosophila)的精子尾巴非常長,有種學名叫作 Drosophila bifurca 的二裂果蠅,擁有記錄上最長的精子,尺寸大到要以公分計算,一隻精子從頭到尾可以長達「5.8 公分」[1],屌打所有動物!
(y編按:人類的精子長度頭部約是3.1~5.1 µm,尾部為50 µm 左右。0.000 1 公分(cm)= 1微米(µm),再想想果蠅跟人類的大小差異,就知道 5.8公分的精子有多巨大了。)
果蠅,為什麼你的精子這麼大?
儘管一般會在垃圾桶附近飛行的那個果蠅,跟 D. bifurca 是不同品種,不過兩者體型上相去不遠,5.8 公分的精子,是它們身長的 20 多倍。這長度是什麼概念呢?以人類來說,假如一位人類男生也能製造這款大小的精子,那麼一隻精子將有十幾層樓那麼高,相當於送人上太空的火箭那麼長,簡直天空才是極限。

製造精子需要耗費男生寶貴的能量,D. bifurca 小小的身軀要製造這麼驚人的精子,花費的代價高昂無比。最常見的果蠅是黃果蠅(Drosophila melanogaster),精子長度是 0.187 公分(其實也不短,足足有人類 300 倍以上),一隻果蠅男生平均一天可以製造 1745 隻精子,相比之下,D. bifurca 的精子長度遠遠超過黃果蠅,一天卻只能生產 221 隻[2]。
男生製造精子,目標是要跟卵結合,把 DNA 傳遞下去。某些果蠅男生為什麼不採取精子海戰術,用相對低廉的成本生產一大堆精子,增加受精的機會,卻要採取這種耗費高昂成本,只能製造少量巨型精子的策略?演化學家將這個疑問稱為「大精子悖論(big sperm paradox)」,是什麼奇妙的理由使果蠅演化出這麼誇張的雄性特徵?
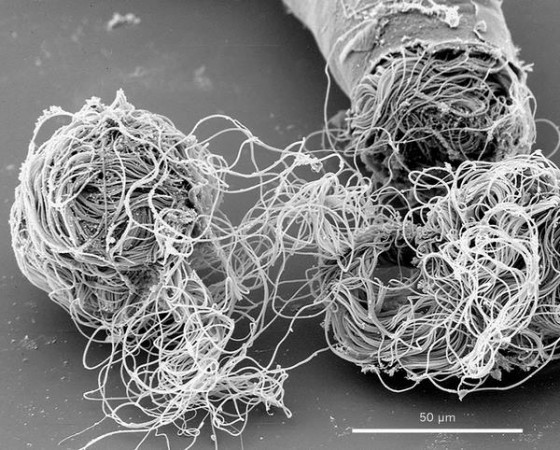
交配後,看不見的生死格鬥才剛開始
男果蠅為了與女生交配,在性擇(sexual selection)中勝出,有套繁複的求偶過程,讀者可以從〈調控多巴胺神經元,讓老齡雄果蠅找回愛愛的動力〉與〈公果蠅追女友竟被她狠心阻止! 朱麗安的雷射追蹤系統〉中稍加體會。然而這兩篇文章提到的都是果蠅生殖行為的前半,也就是「生殖前」的部分。事實上,交配結束後,也就是「生殖後」的競爭(post-copulatory sexual selection),重要性完全不遜於前者。
上半場結束,男女果蠅交配完,男生讓精子進入女體以後,漫長的下半場比賽才剛開始。果蠅的精子並不會直接抵達卵細胞,而是先儲存在附近的器官「儲精囊(seminal receptacle)」等候與卵結合[3]。假如女生後來又跟別的男生交配,不同男生的精子,會在女生的體內搏命競爭,爭取成功受精的機會[4]。這個狀況下,精子的「高品質」有助於在競爭中脫穎而出,成為「幸運的精子」。
對生物來說,把遺傳血脈傳承下去是重要的任務。包括果蠅在內,很多動物的男生為了獲得與女生生寶寶的機會,不惜血本使出像是孔雀開屏、藍寶堅尼、鹿角決鬥等各式花招,無所不用其極,然而以上都是在生殖前的階段,表面上的明爭。許多種動物在交配以後,不同男生精子在女生體內搏出位,如 NBA 勇士隊的 Draymond Green 與雷霆隊的 Steven Adams 般的激烈對抗更是暗潮洶湧。
這檯面下看不見的暗鬥就是「精子競爭(sperm competition)」及「隱性雌選擇(cryptic female choice)」,常常是場慘烈的死鬥,各種鬥爭方式的創意與戲劇性絲毫不輸莎士比亞。例如多種果蠅,先交配的男生雖然可以讓自己的精子先進入女生體內,有望搶得先機,隨後其他男生送入的精子卻又會取代比較早的,這下反而是晚到的有利;還有些動物的男性生殖器配備特殊構造,在交配時可以把之前的精子刮掉;甚至還有的精子主打化學戰,能分泌毒素消滅對手。
愈大愈好!
大尺碼精子有什麼優勢,才能在演化戰爭中勝出?由於隱性雌選擇是檯面下的暗鬥,要闡明其中奧妙並不容易。最近有個研究採用果蠅界最常用的對象黃果蠅,遺傳上小有差異的多個品系,進行同種間的交配實驗,比較精子長度、儲精囊長度、女生再度交配的天數、精子取代率(早到的精子被後來的替換)等因素間的關係[5]。
結果發現,儲精囊的長度與精子長度、精子取代率正相關,卻與再度交配的天數負相關。簡單來講,女生的儲精囊愈長,交配次數愈多,精子長度也愈長,被替換的機率愈大。
這層關係不難解釋,因為精子的長度與數目無法兩全,男生要製造較長的精子,往往沒辦法兼顧數量,女生也就要與不同男生交配更多次,才能獲得足夠的精子儲藏量。也許狀況是,女生演化出愈長的儲精囊,驅使男生的精子也演化得更大,引發一連串互相適應,才造成我們見到的驚人尺寸。

性擇偏好大精子,讓女生間接受益
然而那些果蠅品種中,男生被苦苦相逼製造特大精子,女生真的能從中獲取遺傳利益嗎?果蠅能製造的精子長度不受外界狀況影響,不管環境好壞,精子長度沒有什麼差別,但能生產的精子數目仍受到生長狀況影響,好環境下的男生頭好壯壯,精子不會比較大隻,卻能產出比較多。
偏好大精子對女生有何好處?經過一系列同種內與跨種間的比較與分析後,論文指出,一種果蠅的精子長度較短(例如黃果蠅),意謂製造時耗費資源較少,即使是資質普通的男生也能大量生產,女生難以分辨優劣;若是一種果蠅的精子是大尺寸,品質欠佳的男生負擔不起太多,唯有天賦異稟者才能大批生產,搭配更高的交配次數與精子汰換率,女生便能藉此增加生下優秀男生的寶寶的機率[6] [7]。
既然驅使男生製造大尺寸精子,對女生有這等利益,為什麼我們沒有觀察到全部種類果蠅的精子都愈大愈好?或許是策略不同所致。畢竟演化要付出代價,儲精囊較長與交配次數更多,都會帶給女生負面影響,甚至折損壽命。我們不難想像各種影響因素,最終導致不同果蠅演化出各有巧妙的多變策略,然而一大困難在於,從結果看來可知確有隱性雌選擇發生,然而過程細節究竟如何很難釐清,所以仍有很多地方有待研究。
不論如何,這個新研究非常漂亮地解釋,演化出現特大精子的理由。

看不見的第二性徵—精子
可能有人會認為,頭部的遺傳物質才是精子真正重要的部分,尾巴那麼長根本不實用,是無意義的浪費,然而這正是性擇運作的模式。看看自然界中,眾多誇張的第二性徵,華麗炫耀的羽毛、繁複至極的舞蹈、求婚的 101 朵玫瑰……(好吧,最後一個不太算),這就是性擇的代價,男生用華而不實、過度浪費的炫富,向女生表達「我負擔得起」,換取傳宗接代的機會。
對果蠅精子的研究,拓展了我們對性擇的視野,性擇的方式不只跳舞打架,第二性徵未必是體外的華麗裝飾;性擇也可以是發生在女生體內的精子搏鬥,精子尾巴亦可作為炫耀的指標。男生在生殖後競爭的致勝之道,可能不單純是生產「最多」、或是打造「最好」的精子,而是在「女生體內的特定條件下,能夠最大化存活與競爭力」的精子。
參考文獻:
- Pitnick, S., Spicer, G. S., & Markow, T. A. (1995). How long is a giant sperm. Nature, 375(6527), 109-109.
- Bjork, A., & Pitnick, S. (2006). Intensity of sexual selection along the anisogamy–isogamy continuum. Nature, 441(7094), 742-745.
- Price, C. S., Dyer, K. A., & Coyne, J. A. (1999). Sperm competition between Drosophila males involves both displacement and incapacitation. Nature, 400(6743), 449-452.
- Drosophila Sperm within the Seminal Receptacle viewed under DIC(影片)
- Lüpold, S., Manier, M. K., Puniamoorthy, N., Schoff, C., Starmer, W. T., Luepold, S. H. B., … & Pitnick, S. (2016). How sexual selection can drive the evolution of costly sperm ornamentation. Nature, 533(7604), 535-538.
- Forget peacock tails, fruit fly sperm tails are the most extreme ornaments
- Why fruit fly sperm are giant
本文亦刊載於作者部落格《盲眼的尼安德塔石匠》暨其 facebook 同名專頁。