




教科書與科普書籍常描述向日葵的追日行為雖與刺激方向有關,但不屬於生長,而是膨壓變化所造成,故屬於傾性反應。這樣歸類是否合理?什麼是追日行為?對其運動機制的研究又有什麼新發現?

爭議在哪裡?
大多數動物具有顯著的神經系統與肌肉,遇逆境可立即逃避。
植物運動能力較不顯著,但對環境變化亦可適當反應,包含向性 (tropism) 與傾性 (nasticmovements) 反應。
高中生物教科書對向性的描述如下:
- 「有些刺激會引起植物組織的生長不均勻,而造成和刺激方向有關的運動」
- 「植物受環境刺激時,因局部生長的速率或方向產生改變,導致植物體兩側生長差異」
- 「植物器官因應外界的光照、水分、接觸或地心引力等刺激,使植物組織的生長不均勻,產生和刺激方向有關的生長反應」
大學普通生物學也有類似的內容:
「任何一種導致植物器官朝向或遠離刺激而彎曲生長的生長回應皆稱為向性。」 (Campbell and Reece, 2005)

所以教師與學生常認為「向性」需符合兩項條件:和刺激方向有關的運動,以及肇因於生長不均勻,故向性是種生長現象。
而傾性反應自然就歸類於和刺激方向無關的運動,且與生長無關的現象。基於上述推論,植物的器官於白天追隨太陽東起西落的運動雖與刺激方向有關,但不屬於生長(而是膨壓變化造成),故屬於傾性反應。植物器官追隨太陽東起西落的運動稱為追日行為,雖然現在高中教科書已不再介紹此例,但許多參考書、講義、考題與教師上課的內容中,追日行為始終被歸屬於傾性反應的範疇中。這樣的歸類是否合理?
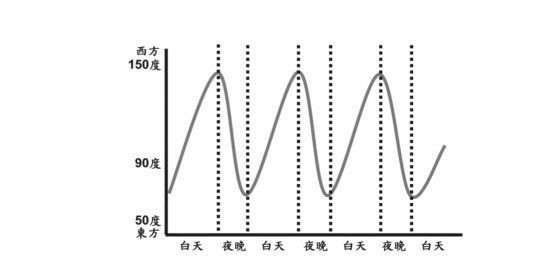
那麼追日行為到底是?
追日行為又稱向日性 (heliotropism) ,源自希臘文 helios (意指太陽,希臘神話中的太陽神即為 Helius )與 tropos(意指轉彎)。
追日行為見於菊科、豆科、錦葵科等 (Ehleringer and Forseth, 1980),其中最為人所知的是向日葵 (Helianthus annuus) ,其葉子、頂端芽和發育中的花序,都表現向日性,其中發育中的花序表現最強的追日行為,在花成熟的過程中其向日性逐漸減緩直到開花,成熟的花朵保持面向東方直到衰老凋萎 (Vandenbrink, et al., 2014) 。

雖然向日葵花序的向日性廣為人知,但科學家多是研究其葉子的向日性機制,對花的向日性機制所知甚少。向日葵幼苗亦具有向光性,其下胚軸受單側光刺激後引起彎曲生長,而莖頂的追日行為直到後來的發育階段才表現,代表兩者具有不同機制 (Vandenbrink, et al., 2014) 。向日性可分為橫向日性 (diaheliotropic) 與側向日性 (paraheliotropism) (Silvertown, and Gordon, 1989; Vandenbrink, et al., 2014) 。
橫向日性使葉子在白天時與太陽光方向垂直,可增加光合作用效率,而側向日性使葉子與太陽光方向平行,以減少強光的傷害。
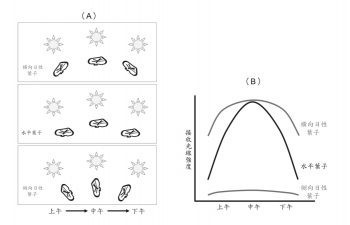
一種「向性」,各自表述
向日性的字根為 -tropism,可知向日性應屬於向性,而與部份教材的歸類不同。
《植物生理學》 (Hopkins and Hüner, 2013) 與《現代生物學》 (Postlethwait and Hopson, 2007) 對「向性」的說明,是指與刺激方向有關的運動,與其是否屬於生長無關。這樣的定義也見於國際學術期刊的研究報告或回顧性文章 (Chen, et al., 2012; Lee, et al., 2014; Silvertown and Gordon, 1989) 。
換句話說,某項植物行為是否屬於向性,與其「是否屬於生長」無關。
葉子的向日性可透過葉柄基部的運動構造:葉枕 (pulvinus) 的膨壓變化所引發。葉枕驅動的運動速度可以很快,例如錦葵科花葵屬的副瓣對消錦葵 (Lavatera cretica) ,其葉子的向日性運動可達每小時偏轉 40 度角。另一方面,向日性也可能是因生長所引發,而不一定是透過細胞的膨壓變化,例如許多具追日性的植物器官不具葉枕,尤其是莖與花,他們透過局部組織生長的方式產生追日行為,但此種生長屬於不可逆的細胞擴展 (Vandenbrink, et al., 2014),與引發向光性的生長方式類似,但此領域的研究極少。

向日性的感光構造為何?對向日葵莖的向日性研究發現其感光受器可能為葉,當去除幼葉(長度小於四公分)後,於數天內可抑制其追日行為;但若去除成熟葉(長度大於四公分)則使追日行為消失,直到幼葉發育成熟
(Vandenbrink, et al., 2014) ,但葉子是否確實為感光受器仍須進一步研究。除了葉子之外,科學家也發現具向日性的高山植物草玉梅 (Anemone rivularis) 花具有追日行為,其花瓣為感光受器 (Zhang, et al., 2010);而雪毛茛 (Ranunculus adoneus) 的感光受器位於莖的頂端,也就是總花梗 (Sherry and Galen, 1998) 。
破除「追日」的刻板印象
◎「是否屬於不可逆的生長」並不是判斷向性的條件。
◎向日葵的追日行為(向日性),其反應方向與刺激方向有關,應屬向性。
◎部分植物器官的追日行為可能屬於生長現象。
情資來源:
- Campbell, N. A. and Reece, J. B. 2005. 生物學第六版。(鍾楊聰等人譯)。臺北市:偉明圖書有限公司。
- Chen, J., Moreau, C., Liu, Y., Kawaguchi, M., Hofer, J., Ellis, N., and Chen, R. 2012. Conserved genetic determinant of motor organ identity in Medicago truncatula and related legumes. Proc. Natl. Acad. Sci. U.S.A. 109(29):11723-11728.
- Ehleringer, J. and Forseth, I. 1980. Solar tracking by plants. Science. 210(4474):1094-8.
- Hopkins, W. G. and N. P. A. and Huner. 植物生理學第四版。(徐善德、廖玉琬編譯)。臺北市:偉明圖書有限公司。
- Lee, H., Garrett, W. M., Sullivan, J., Forseth, I. and Natarajan, S.S. 2014. Proteomic analysis of the pulvinus, a heliotropic tissue, in Glycine max. Int. J. Plant Biol. 5(1):8-12
- Postlethwait, J. H. and Hopson, J. L.2007. Modern Biology Chapter 31: Plant Responses. Holt, Rinehart and Winston.
- Sherry, R. A. and Galen, C., 1998 The mechanism of floral heliotropism in the snow buttercup, Ranunculus adoneus. Plant Cell Environ. 21: 983-993.
- Silvertown, J. and Gordon, D. M. 1989. A Framework for Plant Behavior. Annu. Rev. Ecol. Syst. 20:349-366.
- Vandenbrink, J. P.,Brown, E.A., Harmer, S. L. and Blackman, B. K. 2014. Turning heads: The biology of solar tracking in sunflower. Plant Sci. 224:20-26.
- Zhang, S., Ai, H-L., Yu, W-B., Wang, H. and Li. D-Z. 2010. Flower heliotropism of Anemone rivularis (Ranunculaceae) in the Himalayas: effects on floral temperature and reproductive fitness. Plant Ecol. 209(2):301-312.

本文摘自《生物學學理解碼:從研究史、生態、生理到分子生物,完整剖析39個高中生物學疑難案例》,2019 年 3 月,紅樹林出版。






























{kind=link}