

植物與昆蟲有著緊密的關係,有時兩者是相互依賴、雙方獲利的生命共同體,有時彼此卻是互相競爭、視為敵軍的掠食關係。為了在這場生存遊戲中活下來,刺探敵情、升級軍備當然少不了。不過刺探軍情、升級軍備不是短時間內可完成, 需要漫長的時間演化。
植物的防禦機制
植物與昆蟲間「共演化 (co-evolution) 」的現象,已有數百萬年,兩者在共同演化的過程會互相影響演化的方向及過程。不管在分子生物學、形態學、生物化學上,植物皆已發展出各種防禦機制來應變昆蟲的攻擊。例如:葉片形狀會影響昆蟲的產卵的選擇。在兩片葉片營養價值相同情況時,捲葉象鼻蟲 (leaf-rolling weevil, Apoderus praecellens) 相對較不偏好在有隆起的葉片的產卵及捲葉[1]。從分子學的層面上,植物會藉由激素 (phytohormones)、轉錄因子 (transcription factors) 等訊息傳遞路徑,及防禦蛋白質 (defense proteins)、物理屏障 (physical barriers) 來抵禦食草性昆蟲[2]。這些防禦反應還可因發生的時間點、範圍及位置而有所不同,可直接或是間接抑制草食性昆蟲的能力[3]。其中,就屬有毒性的次級代謝物 (toxic secondary metabolites) 是植物最廣泛且有效的防禦武器。

最有效且最大量的防禦武器——次級代謝物「酚苷」
植物所產生的次級代謝物中,數量最多的就屬酚苷 (phenolic glycosides),這種化學分子可顯著地影響昆蟲的生長及發育,甚至是影響昆蟲的行為[4]。在植物體中,丙二酰化反應 (malonylation) 是修飾酚苷的重要步驟,這個反應是藉由 PMAT (Phenolic glucoside malonyltransferase) 酵素的催化來將丙二酰基 (malonyl group) 轉移至酚苷,藉此代謝毒素 (detoxification)[5]。
農作物的毀滅者——害蟲大軍粉蝨
甘薯粉蝨 (sweet potato whitefly, Bemisia tabaci (Gennadius)) 是個具有強大毀滅性的農作物害蟲,破壞範圍遍及全球,害蟲粉蝨會嚴重地減少農作物的產量,並會咬傷植物韌皮部、傳遞病毒、分泌黏液 (honeydew)。粉蝨會造成大量農作物損傷的原因有二:第一,因為粉蝨是超過百種黴菌、真菌等病原菌的溫床,這些病原菌會透過粉蝨感染植物。第二個原因則是粉蝨的多食性 (polyphagous),目前已知的粉蝨宿主植物 (host plant) 種數至少達 600 種以上,並且其中有很高比例的植物體內都含有酚苷,這顯示了粉蝨體內可能自有一套解決酚苷毒性的妙招,以防止植物的次級代謝物毒死自己。
為什麼粉蝨不怕次級代謝物酚苷的毒性呢?
害蟲粉蝨的存活秘訣,正巧就是針對次級代謝物中丙二酰化反應,藉由產生可以代謝酚苷的毒素的蛋白質,來防止攝取過量的毒素。在粉蝨蟲的體內,含有 BtPMaT1 基因,這個基因會參與在酚苷毒性代謝的作用,來藉此中和植物產生的毒性次級代謝物。從序列比對的結果發現,與 BtPMaT1 基因最相近的同源基因 (homologs) 竟然是在植物中,並且在節肢動物中沒有發現其他的同源基因。顯示粉蝨蟲中的 BtPMaT1 基因來源可能為植物。
為了進一步地瞭解 BtPMaT1 基因在粉蝨中的功能,研究團隊使用了 RNAi (核糖核酸干擾,RNA interference) 的方式,來靜默 (silence) 粉蝨蟲中 BtPMaT1 基因的表現。藉由將特定的雙股 RNA 片段 (dsRNA) 餵食給粉蝨成蟲,使這段 dsRNA 進入粉蝨體內,與 BtPMaT1 基因結合,使其不能順利作用,進而靜默粉蝨中該基因表現。實驗結果顯示,當粉蝨中 BtPMaT1 基因被靜默時,粉蝨成蟲對於酚苷毒素的抵抗力會降低許多,進而提高粉蝨的致死率。
這項研究結果顯示,BtPMaT1 基因在粉蝨中和酚苷毒性的過程中,扮演著重要的角色。這代表了生物學家可以利用一種新方法來處理害蟲粉蝨,且這項方法具有針對粉蝨的專一性,可以完全驅離粉蝨,但不影響其他具有經濟效益的昆蟲。
植物跟昆蟲間的戰役,battle!

要驗證一個基因從兩個物種間平行轉移是相對困難的。因此研究團隊藉由分析植物中與粉蝨 BtPMaT1 相似的基因序列,來了解粉蝨 BtPMaT1 與植物基因有親屬關係 (kin)。除此之外,從其他的親緣分析 (phylogenetic analysis) 的結果,可發現粉蝨的 BtPMaT1 蛋白與植物中 BAHD acyltransferases 群聚 (cluster) 在一起[8],而從 Independent genomic analyses 分析結果,則發現 BtPMaT1 這個從植物轉移來的基因已經完全融入粉蝨本身的基因,再加上 BtPMaT1 基因是在粉蝨成蟲 (adult) 時有較高的表現量,而在卵 (egg) 的階段並不太表現,因此可確認粉蝨蟲中含有 BtPMaT1 基因並不是因為基因污染 (gene contamination) 。依目前的研究結果推論,BtPMaT1 已從植物中平行轉移至粉蝨。
基因在不同物種間流轉早有前例呢
數百萬年以來,植物及昆蟲就已開始從微生物中竊取基因,以及藉由微生物們獲取基因,為的就是後續發展防禦或進攻策略。例如:咖啡果小蠹 (Hypothenemus hampei) 就藉由掠奪微生物的基因,來消化難以分解的細胞壁,藉此攝取更多的養分[6]。小麥也藉由偷取真菌的基因來防治小麥赤黴菌[7]。真菌和微生物會互相擷取基因在早先已被發現,但植物與昆蟲間會互相竊取基因的例子,粉蝨則為首例。從植物平行轉移至粉蝨的 BtPMaT1 基因,其可轉譯出酚苷丙二酰化酵素 (phenolic glucoside malonyl-transferase) ,催化酚苷代謝。使粉蝨可以在體內進行酚苷丙二酰化,藉此中和植物的次級代謝物酚苷(圖一)[8]。
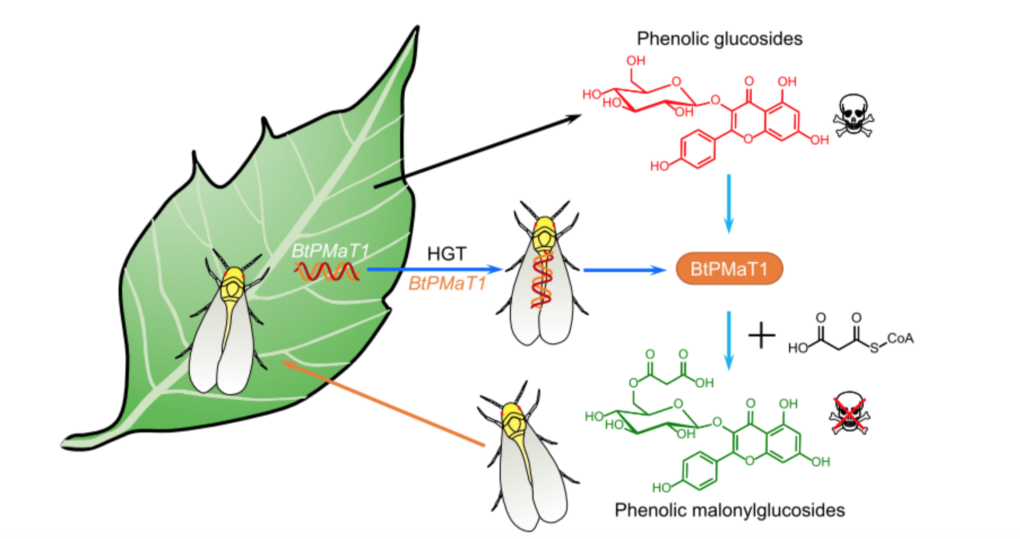
粉蝨是如何竊取植物的基因的呢?
目前仍是未知。生物學家根據現有證據推測,病毒可能可以作為中繼站來運送基因,藉此將植物基因搭載到粉蝨體內。植物與昆蟲間的軍備戰,是隨時在進行的。粉蝨藉由劫取植物代謝毒素的基因來抵擋植物的防禦機制是一場精彩的例證。為了生存,競爭最大利益,植物跟昆蟲今日可能也正在共同演化出更適合當今環境的生存妙招。
參考資料
- Higuchi, Y., & Kawakita, A. (2019). Leaf shape deters plant processing by an herbivorous weevil. Nature plants, 5(9), 959-964.
- Erb, M., & Reymond, P. (2019). Molecular interactions between plants and insect herbivores. Annual Review of Plant Biology, 70, 527-557. [3] Mithöfer, A., & Boland, W. (2012). Plant defense against herbivores: chemical aspects. Annual review of plant biology, 63, 431-450.
- Boeckler, G. A., Gershenzon, J., & Unsicker, S. B. (2011). Phenolic glycosides of the Salicaceae and their role as anti-herbivore defenses. Phytochemistry, 72(13), 1497-1509. [5] Taguchi, G., Ubukata, T., Nozue, H., Kobayashi, Y., Takahi, M., Yamamoto, H., & Hayashida, N. (2010). Malonylation is a key reaction in the metabolism of xenobiotic phenolic glucosides in Arabidopsis and tobacco. The Plant Journal, 63(6), 1031-1041.
- Acuña, R., Padilla, B. E., Flórez-Ramos, C. P., Rubio, J. D., Herrera, J. C., Benavides, P., … & Rose, J. K. (2012). Adaptive horizontal transfer of a bacterial gene to an invasive insect pest of coffee. Proceedings of the national academy of sciences, 109(11), 4197-4202.
- Wang, H., Sun, S., Ge, W., Zhao, L., Hou, B., Wang, K., … & Kong, L. (2020). Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science, 368(6493).
- Xia, J., Guo, Z., Yang, Z., Han, H., Wang, S., Xu, H., … & Zhang, Y. (2021). Whitefly hijacks a plant detoxification gene that neutralizes plant toxins. Cell.
- nature-First known gene transfer from plant to insect identified


































《運動基因》立體封面72dpi.jpg)

