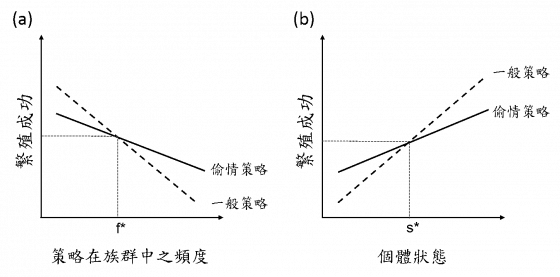- 文字編輯/翁郁涵
在〈偷情也是一種選擇?小王老王是怎麼來的?──動物的另類繁殖策略(上)〉,我們已經認識了動物們的「偷情花招」和為何夫妻們與小王老王之間會有這麼多的「情慾流動」,接下來,讓我們來看看幾種「花名在外」的動物吧!
命中注定我偷情:替代型另類繁殖策略
由基因所決定的替代型另類繁殖策略在動物界中相當稀少,其中最著名的案例是側斑蜥 (side-blotched lizard,Uta stansburiana) 與流蘇鷸 (ruff,Philomachus pugnax)。

(a) 側斑蜥的三種繁殖策略與喉部顏色:橘喉雄蜥為占領大領地的領域雄蜥,藍喉雄蜥為維持小領地的領域雄蜥,黃喉者為無領域性的偷情者。
(b) 流蘇鷸的三種雄鳥與雌鳥:上為雌鳥,下排由左至右分別為領域雄鳥、費德雄鳥與衛星雄鳥。
圖/作者整理 圖片來源:a: Sinervo Lab 網站;b: Farrell et al, 2013
採取不同策略的側斑蜥雄蜥,會對應三種不同的喉部顏色(上圖a),分別是兇猛強勢的橘喉好鬥者、穩定禦地的藍喉防衛者及無領域的黃喉偷情者,三種策略且剛好互相剋制,橘剋藍、藍剋黃、黃剋橘,形成剪刀石頭布的勝負循環。目前側斑蜥的繁殖策略已經被證實是由單一基因座的三個等位基因所調控3。
流蘇鷸的雄鳥同樣有三種不同的繁殖策略:領域雄鳥(independent)、衛星雄鳥(satellite)與費德雄鳥(feader),前者體型最大,行守護領域的繁殖策略,占有多數雌鳥,後二者則都行偷情策略(上圖b)。衛星雄鳥體型稍小於領域雄鳥,游移在領域雄鳥周圍等待機會;費德雄鳥則直接擬態成雌性,混在領域雄鳥的後宮中。
流蘇鷸的三種行為都是基因決定,領域與衛星行為由一對等位基因調控,費德雄鳥的基因則獨立於此基因之外,與體色相關的 MC1R(沒錯,就是 X 教授拿來撩妹的基因!)相互關聯4。
一切都是命運的決定:情況決定型另類繁殖策略
情況決定型的另類繁殖策略在動物界則相當常見,昆蟲、魚類、兩生類、爬行類、鳥類、哺乳類都有不少研究案例。

(c) 嗡蜣螂屬,Onthophagus nigricornis 雄蟲。
(d) 藍鰓太陽魚的三型雄魚與雌魚:左上為雌性擬態型雄魚,正中體型最大者為親護型雄魚,其下方為雌魚,體型最小的鬼祟型雄魚在最下方。
圖/作者整理 圖片來源:c: Wikiwand;d: Neff BD & Svensson EI, 2013
無脊椎動物中最經典的例子莫過於嗡蜣螂屬物種 (Onthophagus spp.,糞金龜的一屬) 的犄角型態與對應的繁殖策略(上圖c)。其雄蟲有具角與無角兩種型態,具角雄蟲體型大,利用犄角打鬥以爭奪雌性,無角雄性體型較小,只好偷情者策略。實驗發現幼蟲期的飲食情況將直接影響成蟲的型態與策略,吃得好的大體型幼體在化蛹後會發育成長角好鬥的雄蟲,體型小的幼體則會發育為無角的偷情雄性5。
脊椎動物中不能錯過的物種,則是屬於時序型繁殖策略轉換的藍鰓太陽魚 (Lepomis macrochirus)。藍鰓太陽魚有三種雄性,分別是體型最大的親護型 (parental)、體型最小的鬼祟型 (sneaker) 與體型中等的雌性擬態型 (female mimic),後二者都是偷情式繁殖策略。雄性幼魚在第二年時會決定發育成親護型或是偷情型,若是親護型,則需要再等三至五年才能性成熟,偷情路線則是一開始就成為鬼祟型的小雄魚,在親護型領域附近打游擊戰,兩三年後體型漸大,即轉變成雌性擬態型,進入親護型雄性的領域內,偷偷與真正的雌性交配(上圖 d)2。
小王老王或元配,雌性會怎麼選?
與偷情者交配有好處也有壞處6,如果利大於弊,雌性出軌傾向將會大大增加;反之,若弊大於利,則會排斥偷情者的追求。
出軌最直接的壞處是被抓猴以後,原配雄性對子代投注的心力會下降。在動物界中,雄性的直覺不比雌性差,尤其是對這種綠光罩頂的情況。
許多雄性能夠經由各種訊息來判斷雌性是否出軌,再調整親代照護的能量投資,進而影響子代存活率,這對偷情的雌魚來說將是很大的損失。另一壞處是偷情者的行為會對卵造成擾動,將會降低受精卵的存活率,這對體外受精的類群有非常大的影響。

至於出軌的好處,是受精率的提升,以及子代基因多樣性的提高。每個卵都需要投資相當多的資源,對雌性來說,將卵排出體外卻未受精是極度浪費的,所以有越多雄性的精子來提升受精率是很重要的。另外,與多種雄性交配能夠讓子代維持基因多樣性,在基因層面上分散風險,以免所有子代在日後一次陣亡。
最後,雌性需要評估的是偷情者與原配的基因品質,若偷情者基因品質很高(勇猛帥氣的小鮮肉或老王),則對子代的基因有利,反之則有弊無利。綜合考量這些利弊後,雌性將會對偷情者顯示出不同程度的偏好,讓配對雄性有不同程度的戴綠帽機會。當然,偷情者不一定會在意雌性的偏好,許多類群的偷情者會用強迫的方式完成交配或受精行為,讓人防不勝防。
無論正規非正規,都是為了傳遞基因
偷情策略在動物界相當常見,少數物種的偷情策略是由基因決定的替代型,命中注定要當個壞男人,不過多數例子是情況決定的固定型或彈性型策略,因應自身與環境情況來決定當個好男人或是小王老王。對地方媽媽來說,需要評估的是受精率上升與基因多樣化的好處,抓猴後親代照護減少的壞處,以及偷情者與原配的基因品質。
無論是雄性或雌性,採取正規或非正規的繁殖策略,其實都只是為了順利傳遞自己的基因。以後各位看到綠綠的社會新聞,甚至是親身體驗的時候,可以回過頭來看看這些大自然的例子,感受一下演化的美妙之處。
參考資料
- Neff BD and Svensson EI (2013) Polyandry and alternative mating tactics. Trans. R. Soc. B 368. 20120045
- Taborsky M and Brockmann HJ (2010). Alternative reproductive tactics and life history phenotypes. In Animal behaviour: evolution and mechanisms (pp. 537-586). Springer, Berlin, Heidelberg
- Corl A, Davis AR, Kuchta SR, and Sinervo B (2010) Selective loss of polymorphic mating types is associated with rapid phenotypic evolution during morphic speciation. PNAS 107, 4254-4259
- Farrell LL, Burke T, Slate J, McRae SB and Lank DB (2013). Genetic mapping of the female mimic morph locus in the ruff. BMC genetics 14, 109
- Emlen DJ (1994) Environmental control of horn length dimorphism in the beetle Onthophagus acuminatus (Coleoptera: Scarabaeidae). R. Soc. Lond. B 256, 131-136.
- Reichard M, Le Comber SC and Smith C (2007). Sneaking from a female perspective. Animal Behaviour 74, 679-688