

文/吳依璇、熊衎昕
櫻花蝦初登場
餓了嗎?來碗櫻花蝦炒飯吧!

來到屏東東港鎮或小琉球就該來吃當地的名產─櫻花蝦!但是為什麼這裡會產櫻花蝦呢?

正櫻蝦,俗稱櫻花蝦,學名Sergia lucens,英文名sergestid shrimp。成蝦身長約40mm,全身佈滿約160個左右發光器及紅色素,呈現半透明紅色。在海中就像是散落吧,千本櫻片片漂流的櫻花而得名,也是馬路東港三寶之一。

櫻花蝦喜歡棲息在水深約150公尺、含有大量泥沙的海底峽谷周遭的海域。日落前上浮,日出後下潛。櫻花蝦吃著比她小的磷蝦們,也被大眼鯛、白帶魚等補食,在大海裡載浮載沉,過著牠的浮游蝦生。
最早,大森教授(Omori ,1969)認為只有在日本靜岡縣駿河灣沿海以及東京灣、相模灣等地才會有這種柔柔亮亮、閃閃動人如同海中落櫻的櫻花蝦。殊不知多年以後,大森教授(Omori, 1988)發現臺灣東港居然也有櫻花蝦!
大森教授是怎麼知道臺灣有櫻花蝦的呢?原來是游祥平教授特地告訴大森教授,大森教授原本還不相信,看到標本以後特地衝到臺灣來鑑定的。
何處是蝦窩?
在臺灣西南海域裡,櫻花蝦最愛窩在高屏和枋寮海底峽谷頭部附近,也就是東港外海(Lee et al., 2004)。
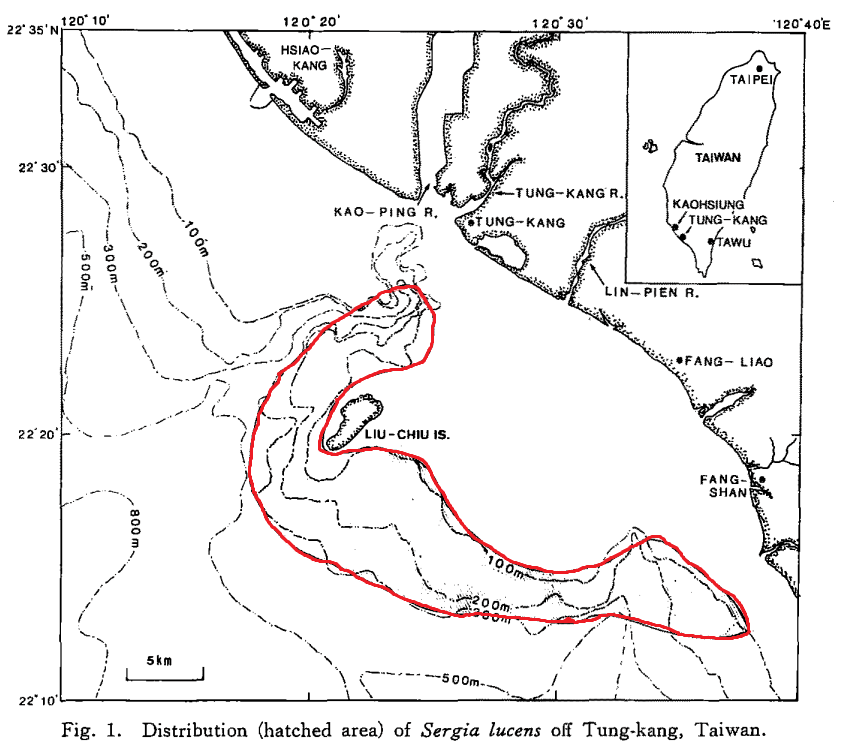
目前櫻花蝦為什麼對生活在高屏和枋寮海底峽谷頭部附近情有獨鍾,至今還沒有明確結論。可能是因為東北側有陸地阻擋了東北季風進入,以及高屏溪、東港溪、林邊溪、率芒溪、枋山溪和楓港溪等河水匯入造成多變的海洋環境(lee et al., 2004),峽谷頭部內複雜的湧升流使底部含有豐富營養鹽的冷水湧到表層(Chiou et al., 2011),尤其高屏溪是把來自陸上富含營養鹽的沈積物直接輸送到高屏海底峽谷(Chiang and Yu, 2010; Chiou et al., 2000)。豐富的營養鹽引來浮游生物,浮游生物則引來櫻花蝦在此聚居。

但是,櫻花蝦不是固定在某一地點生活,受到不同因素影響,春夏季時牠們會在北邊一點,秋冬時則往南邊一點,因此漁場也隨著季節改變(Lee et al., 2004)。

消失的菜餚?
櫻花蝦營養好賣價高,人人見了都喜愛,許多漁民爭先恐後的捕撈櫻花蝦,曾一度造成櫻花蝦產量銳減。為了讓櫻花蝦產業可以永續發展,在「櫻花蝦博士」陳守仁博士的大力協助下,水試所和區漁會於 1992 年號召船主成立櫻花蝦產銷班,並於 1994 年訂定公約及管理辦法,規定每年6月至10月為禁捕期,改捕撈相似種之中型毛蝦。在撈捕期時,每星期大約捕撈 4 天,每年共捕撈 100 天左右,而且要有「櫻花蝦採捕許可牌」的船隻才能撈捕櫻花蝦。產銷班公約還包括每船每日漁獲量和違規相關處罰等規定,希望櫻花蝦能夠有充裕的時間好好長大被人類吃掉。
來看看櫻花蝦船長是怎麼捕捉櫻花蝦的吧,討海人的生活比想像中來得辛苦多了呢。
訂定產銷班公約不只是櫻花蝦漁業資源不但可以永續利用,也可以避免物賤傷農的情況,對於櫻花蝦、漁民和消費者來說是創造雙贏或多贏的局面。人類的欲望早已造成許多魚種面臨滅絕的危機(譬如也是東港三寶之一的黑鮪魚)。多想兩分鐘,我們可以避免濫捕造成物種滅絕,蝦毀人亡。
參考資料:
- Omori, M. (1969). The Biology of a Sergestid Shrimp Sergia Lucens Hansen, Bull. Ocean Res. Inst., Vol. 4, pp. 1-83 (1969).
- Omori, M., Ukishima, Y., and Muranaka, F., (1988). New Record of Occurrence of Sergia Lucens (Hansen) (Crustacean, Sergestidae) off Tung-Kang, Taiwan, with Special Reference to Phylogeny and Distribution of the Species. J. Ocean. Soc., Vol. 44, pp. 261-267.
- Lee, K. T., Liao, C. H., Su, W. C., Hsieh, S. H., Lu, H. J. (2004). THE FISHING GROUND FORMATION OF SERGESTID SHRIMP (Sergia lucens)IN THE COASTAL WATERS OF SOUTHWESTERN TAIWAN. Journal of Marine Science and Technology, Vol. 12, No. 4, pp. 265-272
- Chiou, M. D., Jan, S., Wang, J., Lien, R. C., Chien H. (2011). Sources of baroclinic tidal energy in the Gaoping Submarine Canyon off southwestern Taiwan. JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 116, C12016.
- Chiang C. S., Yu, H. S. (2010). Sedimentary erosive processes and sediment dispersal in Kaoping submarine canyon. Sci China Earth Sci, doi: 10.1007/s11430-010-4076-y.
- Chiou, W. D., Wu, C. C., Cheng, L. Z., (2000). Spatio-temporal distribution of sergestid shrimp Acetes intermedius in the coastal waters of southwestern Taiwan. Fisheries Science. Volume 66, Issue 6, pages 1014–1025, December 2000.
- Wanga, Y.H., Leeb, I.H., Liu, J.T. (2008). Observation of internal tidal currents in the Kaoping Canyon off southwestern Taiwan. Estuarine, Coastal and Shelf Science.Volume 80, Issue 1, 20 October 2008, Pages 153–160.
- 陳守仁.黃建智.陳羿惠.吳龍靜,流動的水晶─臺灣櫻花蝦,科學發展 2010年8月,452期。http://ejournal.stpi.narl.org.tw/NSC_INDEX/Journal/EJ0001/9908/9908-04.pdf。
原刊載於滔滔 Ocean says







































