紅卷柏(File:Selaginella erythropus)是一種耐陰的原始維管束植物。Source: wikipedia
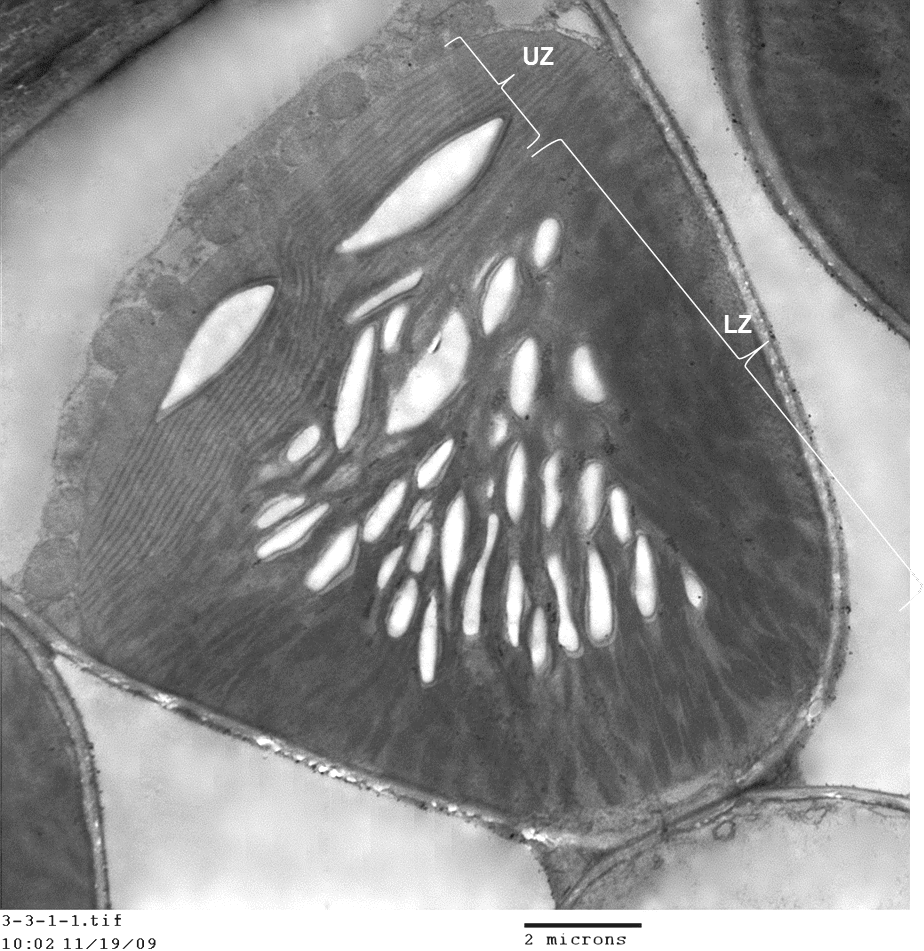
國立中興大學生命科學系副教授許秋容與嘉義大學、中研院、高雄區農改場共組的研究團隊,揭開植物界新形式葉綠體的形成之因,發現紅卷柏的葉綠體內部結構的形成和分裂,具有可塑性,會受光的强度和方向影響而改變,此項葉綠體獨特的發育變異研究成果,提供重要的科學新知和未來新的葉綠體研究方向,研究結果今年4月刊登於知名的《美國植物學期刊 (American Journal of Botany)》。
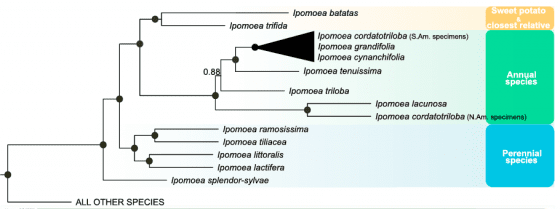
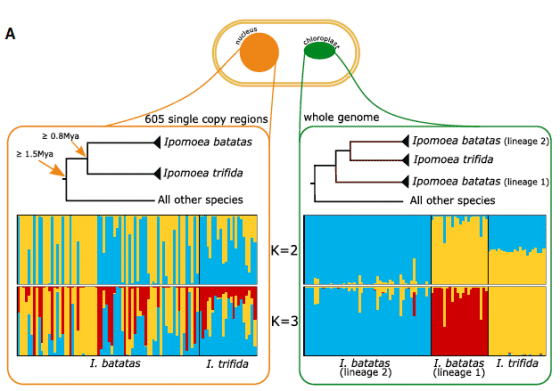
然而,比較親戚間的遺傳差異以後,番薯的六套 DNA 彼此間都更為接近,沒有見到上述狀況。可見番薯的染色體應該只有單一來源,並未經歷過異種合體。家族中所有成員,與番薯最接近的是大星牽牛(Ipomoea trifida, 上上圖B),由此推論,番薯應該是由二倍體的大星牽牛演化而成的「同源多倍體(autopolyploidy)」。
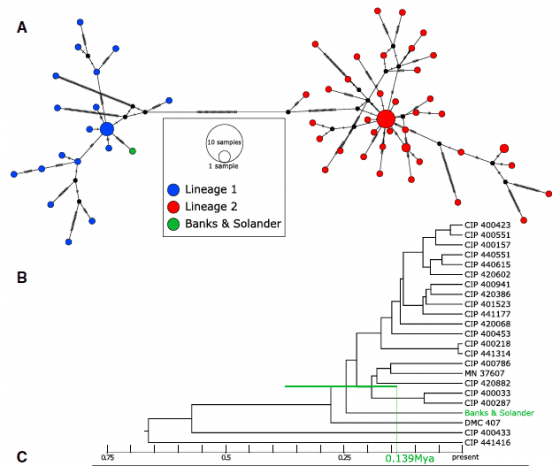
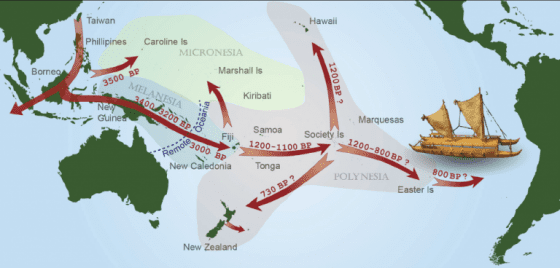
第一部分的質疑點在於,古代番薯樣本是否經過可靠的程序處理?新研究的根據來自古代番薯樣本,這個番薯不是普通的番薯,而是由庫克船長隊伍中的 Joseph Banks 與 Daniel Solander,1769 年採集自社會群島的珍貴標本。定序 DNA 後,得知其葉綠體型號屬於 CLI,估計與美洲親戚至少在 11.5 萬年前分家,遠遠早於人類出現的年代。
1. Mu, P., Carruthers, T., Wood, J. R., Williams, B. R., Weitemier, K., Kronmiller, B., … & Rausher, M. D. (2018). Reconciling Conflicting Phylogenies in the Origin of Sweet Potato and Dispersal to Polynesia. Current Biology, 28(8), 1246-1256.
2. Scotland, R. W., Munoz, P., & Carruthers, T. (2018). Temporal Dynamics of the Origin and Domestication of Sweet Potato and Implications for Dispersal to Polynesia. bioRxiv, 309799.
3. Thomson, V. A., Lebrasseur, O., Austin, J. J., Hunt, T. L., Burney, D. A., Denham, T., … & Linderholm, A. (2014). Using ancient DNA to study the origins and dispersal of ancestral Polynesian chickens across the Pacific. Proceedings of the National Academy of Sciences, 111(13), 4826-4831.
4. Storey, A. A., & Matisoo-Smith, E. A. (2014). No evidence against Polynesian dispersal of chickens to pre-Columbian South America. Proceedings of the National Academy of Sciences of the United States of America, 111(35), E3583.
5. Gonçalves, V. F., Stenderup, J., Rodrigues-Carvalho, C., Silva, H. P., Gonçalves-Dornelas, H., Líryo, A., … & Willerslev, E. (2013). Identification of Polynesian mtDNA haplogroups in remains of Botocudo Amerindians from Brazil. Proceedings of the National Academy of Sciences, 110(16), 6465-6469.
6. Malaspinas, A. S., Lao, O., Schroeder, H., Rasmussen, M., Raghavan, M., Moltke, I., … & Albrechtsen, A. (2014). Two ancient human genomes reveal Polynesian ancestry among the indigenous Botocudos of Brazil. Current Biology, 24(21), R1035-R1037.
7. Moreno-Mayar, J. V., Rasmussen, S., Seguin-Orlando, A., Rasmussen, M., Liang, M., Flåm, S. T., … & Willerslev, E. (2014). Genome-wide ancestry patterns in Rapanui suggest pre-European admixture with Native Americans. Current Biology, 24(21), 2518-2525.
8. Fehren-Schmitz, L., Jarman, C. L., Harkins, K. M., Kayser, M., Popp, B. N., & Skoglund, P. (2017). Genetic Ancestry of Rapanui before and after European Contact. Current Biology, 27(20), 3209-3215.
12. Barbujani, G., Bertorelle, G., & Chikhi, L. (1998). Evidence for Paleolithic and Neolithic gene flow in Europe. American journal of human genetics, 62(2), 488.
生物會受到氣候變遷的影響,氣候也會受到生物活動的影響而發生改變。根據地球系統的能量收支平衡原理,當地表狀態改變時可能同時改變了局地的輻射能量收支平衡與水文循環的機制,再造成局地氣候狀態改變。美國的西北太平洋國家實驗室(Pacific Northwest National Laboratory)近期一項研究成果顯示,人為灌溉所造成的地表植被改變,不只增加土壤中的水分含量,更進一步改變了當地的蒸發與蒸散量,所導致的局地氣候反應是大氣比較容易形成淺雲(shallow clouds)。淺雲的存在會增加當地的反照率(albedo),進一步透過大氣輻射平衡機制導致當地的氣溫下降。